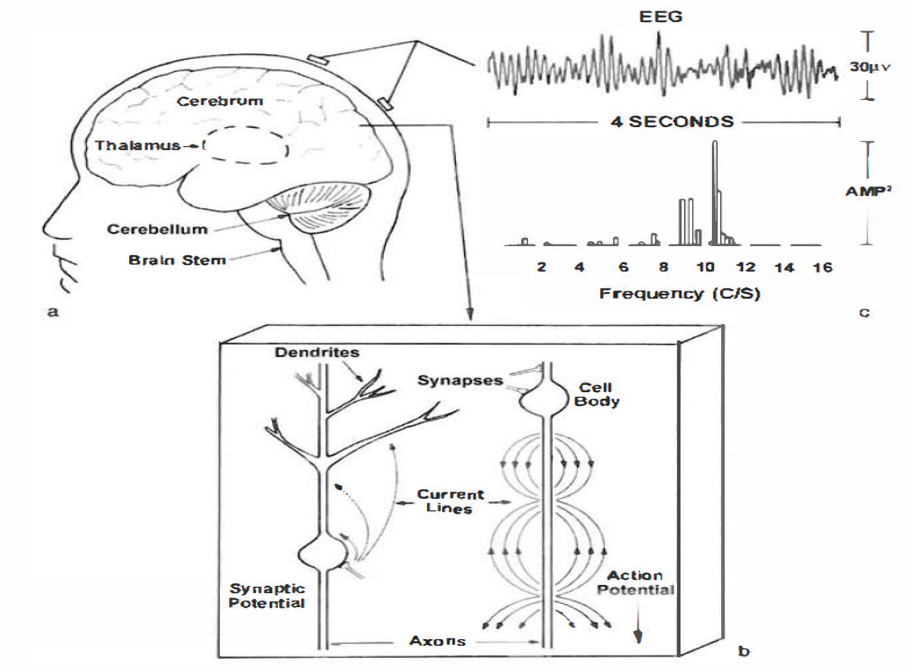
The brain uses a sophisticated communication and command-and-control system that monitors and manages interactions between roughly 100 billion neurons, each with 5,000-10,000 synaptic connections, for as many as 500 trillion synapses in adults (Breedlove & Watson, 2020).
Neuroplasticity, the remodeling of neurons and neural networks with experience, is responsible for learning and memory and makes neurofeedback training possible.
International QEEG Certification Board Blueprint Coverage
This unit addresses II. Basic Neurophysiology and Neuroanatomy - A. Neurophysiology (4 hours).
This unit covers the Bioelectric Origin and Functional Correlates of EEG, Definition of ERPs and SCPs, and Neuroplasticity.
Please click on the podcast icon below to hear a full-length lecture.
Bioelectric Origin and Functional Correlates of EEG
Types of Neurons
We can divide neurons into sensory, motor, and interneurons. Glial cells like astrocytes and microglia work in partnership with neurons.
Sensory neurons are specialized for sensory intake. They are called afferent because they transmit sensory information towards the central
nervous system (brain and spinal cord). The graphic below is courtesy of leavingcertbiology.net.
Motor neurons convey commands to glands, muscles, and other neurons. They
are called efferent because they convey information towards the periphery. Graphic courtesy of leavingcertbiology.net.
Interneurons provide the integration required for decisions, learning and
memory, perception, planning, and movement. They have short processes, analyze incoming information, and distribute their analysis with other neurons in their network. Interneurons are entirely confined to the central nervous system, account for many of its neurons, and comprise most of the brain (Breedlove & Watson, 2020). Local interneurons analyze small amounts of information provided by neighboring neurons. Relay interneurons connect networks of local interneurons from separate regions to enable diverse functions like perception, learning, and memory, and executive functions like planning (Carlson & Birkett, 2021). Graphic courtesy of leavingcertbiology.net.
The
cell body or soma contains the machinery for the neuron’s
life processes. It receives and integrates EPSPs and IPSPs, small graded positive and negative changes in
membrane potential generated by axons. The cell body of a typical neuron is
20 μm in diameter, and its spherical nucleus,
which contains chromosomes comprised of DNA, is 5-10
μm across. The cell body is the only location where neurons manufacture proteins (like enzymes,
receptors, and ion channels) and peptides (neurotransmitters like oxytocin) since this requires ribosomes. Check out the Khan Academy YouTube video, Anatomy of a Neuron.
Dendrites are branched structures designed to receive messages from other neurons via axodendritic synapses (junctions between axons and dendrites shown below) and send messages to other neurons dendrodendritic synapses (junctions between the dendrites of two neurons). Dendrites receive thousands of synaptic contacts and have specialized proteins called receptors for neurotransmitters released into the synaptic cleft (Bear, Connors, & Paradiso, 2020).
A neuron's dendrites are called a dendritic tree, and each branch of the tree is called a dendritic branch. Graphic by BruceBlaus from Wikipedia article Neuron.
An axon is a cylindrical structure only found in neurons that is specialized for the distribution of information within the central and peripheral nervous systems. Axons range from 1 to 25 µm in diameter and 0.1 mm to more than a meter in
length. Over 90% of neurons are interneurons whose axons and dendrites are very short and do not
extend beyond their cell cluster. Axons usually branch repeatedly. Each branch is called an axon collateral.
Axons transmit
action potentials
toward a neuron's terminal buttons. Using microtubules, an axon also bidirectionally transports molecules between the cell body and terminal buttons.
An axon hillock is a swelling of the cell body where the axon
begins. The middle of an axon is the axon proper, and the end is the axon terminal (Bear, Connors, & Paradiso, 2020). Graphic by M.alijar3i from the Wikipedia article Axon Hillock.
The axon hillock sums EPSPs and IPSPs over milliseconds to generate an action potential.
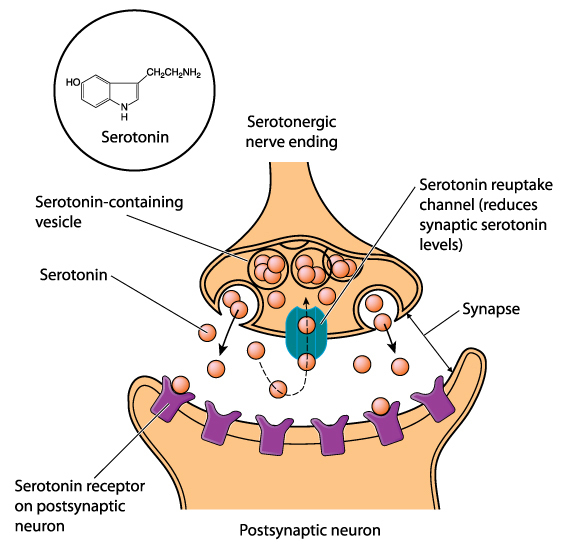
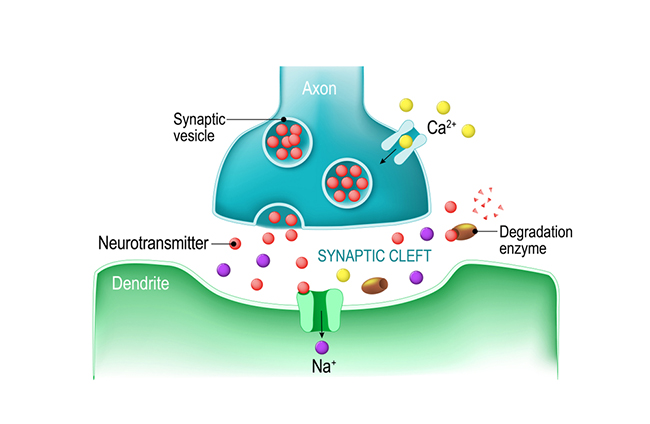
Axon terminals are buds located on the ends of axon branches
that form synapses and release neurochemicals to other neurons. Axon terminals contain vesicles that store
neurotransmitters for release when an action potential arrives. Their presynaptic membrane
may have reuptake transporters that return neurotransmitters from the synapse or extracellular space for
repackaging. The graphic of serotonin reuptake transporters below is courtesy of NIDA.
Types of Glial Cells
While there are hundreds of types of neurons, there are only four main categories of glial cells (astrocytes,
microglia, oligodendrocytes, and Schwann cells).
Old school view: glial cells mainly provide
structural support (glia is derived from the Greek for glue).
New school view: glial cells help
neurons process information, including modulating neuron excitability.
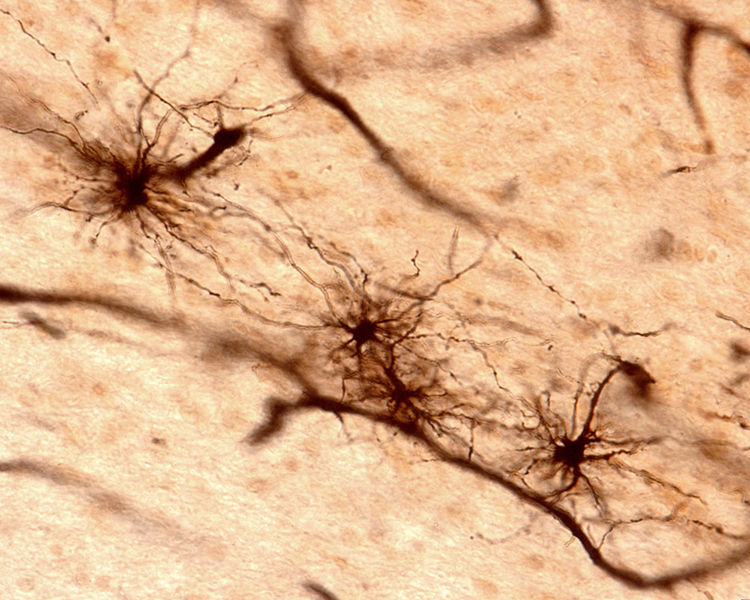
Astrocytes (shown below) are star-shaped and are the most prevalent glial cells in the brain. They guide neuronal migration in the embryo and fetus. Since they occupy most of the expanse between neurons and are separated from neurons by nearly 20 µm, they may affect the growth or retraction of axons and dendrites (collectively called neurites). An emerging view is that astrocytes help neurons process information and communicate with each other through parallel astrocyte-astrocyte networks. Astrocyte membranes contain neurotransmitter receptors that can initiate changes in their membrane potential and internal biochemical processes.
Astrocytes regulate circulating molecules in the extracellular space (the region surrounding neurons) and the synapse. They enclose synapses, limit the movement of released neurotransmitters, and transport them from the synapse to the axon terminal. They also regulate the concentration of ions like potassium outside of neurons to prevent interference with their performance (Bear, Connors, & Paradiso, 2020).
Astrocytes transport nutrients, remove wastes, store glycogen during stage-3 sleep, dynamically control local blood flow, and develop and maintain the blood-brain barrier.
Oligodendrocytes block axonal regeneration by releasing growth inhibitory proteins. These molecules are part of the reason
for minimal functional recovery in the CNS following spinal cord damage. Multiple
sclerosis, a demyelinating disease, destroys oligodendrocytes.
Check out the Blausen Demyelination in the CNS: Multiple Sclerosis animation.
Graded positive and negative changes in membrane potential, called excitatory postsynaptic potentials
and inhibitory postsynaptic potentials, are essential to the EEG and communication among neurons.
An excitatory postsynaptic potential (EPSP) is a subthreshold depolarization that makes the membrane
potential more positive and pushes the neuron towards its excitation threshold. EPSPs are produced when
neurotransmitters bind to receptors and cause positive sodium ions to enter the cell. At a single synapse, a postsynaptic membrane may have tens to thousands of transmitter-gated ion channels. The amount of transmitter released determines how many of these channels will be activated. The size of an EPSP will be a multiple of the number of vesicles, each containing several thousand transmitter molecules. Check out the Blausen Positive Potential animation.
An inhibitory postsynaptic potential (IPSP) is a hyperpolarization that makes the membrane potential
more negative and pushes the neuron away from its excitation threshold. At most inhibitory synapses, IPSPs are produced when
neurotransmitters like GABA or glycine bind to receptors and cause negative chloride ions to
enter the cell. When an inhibitory synapse is closer to the soma than an excitatory synapse, it can counteract positive current flow and decrease the size of the EPSP. This mechanism is called shunting inhibition (Bear, Connors, & Paradiso, 2020).
Integrating Postsynaptic Potentials
Integration is the summation of EPSPs and IPSPs at the unmyelinated axon
hillock.
The axon hillock of a postsynaptic neuron uses two methods to sum EPSPs and IPSPs: spatial and temporal
summation.
In spatial summation, the axon hillock sums the simultaneous postsynaptic potentials (PSPs) from
thousands of synapses on dendrites. In temporal summation, the axon
hillock adds the PSPs from presynaptic neurons that repeatedly fire within a 1-15-ms time window.
An action potential is a brief electrical impulse that transmits
information from the axon hillock to the terminal button. This wave of positive charge only travels in one direction because the preceding segment is refractory due to the closing of its sodium channels. An action potential takes 1-2 ms from the point the
axon hillock reaches its threshold to its repolarization to a negative resting potential.
Action potentials travel down axons, which branch multiple times and terminate at synapses. The all-or-none law and rate laws describe action potential transmission. The all-or-none law states that once an action potential is triggered in an axon, it is propagated, without decrement, to the end of the axon. The rate
law states that neurons represent the intensity of a stimulus by variation in the rate of axon
firing. More intense stimuli shorten the interval before a neuron can fire again, allowing a neuron to fire more
rapidly. An intense stimulus can cause a neuron to fire every 2 or 3 ms, while a weak stimulus might lengthen
the time lag to every 4 or 5 ms.
We can compare action potential conduction to the movement of water through a leaky garden hose.
Garden hose: water can take two paths, inside the hose or through holes in its wall, and the majority of the water will flow where movement is easiest. For a small-diameter hose with many large holes, most of the water will travel through the leaks. Conversely, for a large-diameter hose with only a few small holes, the bulk of the water will remain inside.
This method also consumes considerable energy since sodium-potassium transporters, powered by ATP, are located
across the axon membrane to exchange three sodium for two potassium ions.
Conversely, for a large-diameter axon with few open ion channels, the bulk of the current will remain inside the axon's interior. Wider spacing between adjacent ion channels means that the action potential can depolarize a longer axon segment, which increases conduction velocity (Bear, Connors, & Paradiso, 2020).
Medium-to-large diameter myelinated axons transmit action potentials using a method called saltatory conduction. Each segment of insulating myelin is almost 1-mm
long. The gaps between segments, called nodes of Ranvier,
are 1 to 2 thousandths of a millimeter.
An action potential weakens under each myelinated segment (cable properties) and is then regenerated at each Ranvier node. The destruction of this insulation by demyelinating diseases like multiple sclerosis (MS) can be devastating because it disrupts neuron-to-neuron communication.
Neurons communicate through the release of neurochemicals and ions. Axon
terminal buttons release neurochemicals across a 20-40-nm fluid-filled gap between presynaptic and postsynaptic
structures called
a synaptic cleft and into the extracellular fluid surrounding the neuron.
Chemical synapses produce short-duration (millisecond) and long-duration (seconds to days) changes in the nervous system. Check out the YouTube videos, Neurotransmitter Synapse 3D animation and Neuronal Synapses.
Old-school view: according to Dale’s law, a neuron can only release one neurotransmitter
at a synapse.
New-school view: neurons can release a classical neurotransmitter and a peptide.
Axoaxonic synapses do not affect the generation of an action potential, only the amount of neurotransmitter distributed. In presynaptic facilitation, a neuron increases the
presynaptic neuron's neurotransmitter release by delivering a neurotransmitter that increases calcium ion
entry into its terminal button. In presynaptic inhibition, a neuron decreases
neurotransmitter release by reducing calcium ion entry. These modulatory effects are confined to a single
synapse (Breedlove & Watson, 2020).
Types of Neurotransmitters
While the actual number of neurotransmitters is not known, more than 200 molecules have been identified. Each neurotransmitter may have multiple receptors. A neurotransmitter's effect, excitatory or inhibitory, depends on its interaction with specific receptors. The same neurotransmitter can produce opposite results at different receptor subtypes (Breedlove & Watson, 2020).
The principal neurotransmitter families include amino acid neurotransmitters (GABA, glutamate), amine neurotransmitters (acetylcholine, dopamine serotonin), peptide neurotransmitters, also called neuropeptides (oxytocin, vasopressin), gas neurotransmitters (nitric oxide, carbon dioxide), and lipid neurotransmitters (anandamide and AG-2). The table below is adapted from Breedlove and Watson (2020).
Cholinergic cell bodies and their projections originate in the basal forebrain and brainstem. Cholinergic pathways are involved in arousal, attention, memory, motivation, muscle contraction, and sleep. Watch the Blausen Chemical Synapse: Cholinergic Synapse animation.
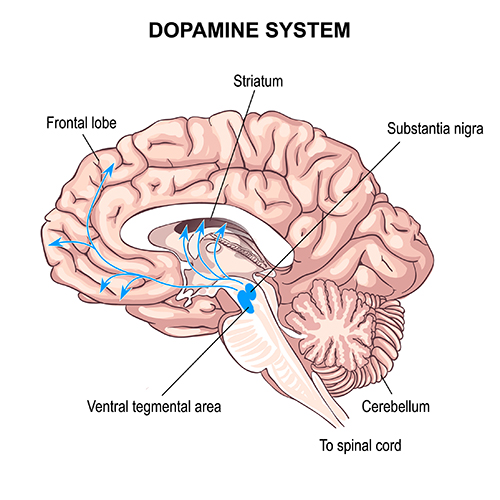
Dopaminergic pathways
Two major dopaminergic pathways
originate in the midbrain: the mesostriatal and mesolimbocortical pathways. Dopaminergic pathways are involved in addiction, motor control, and salience (reward- and threat-based motivation). Check out the Blausen Parkinsons Disease animation.
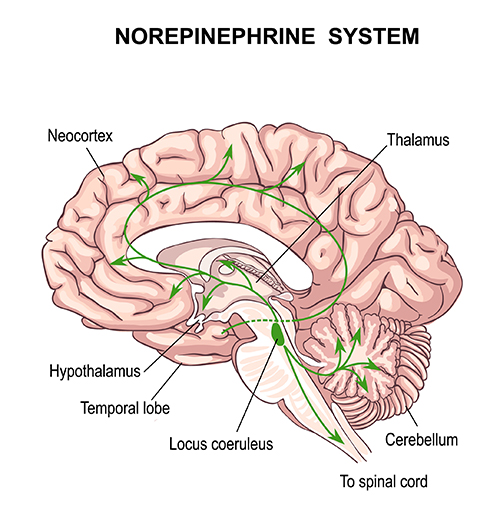
Noradrenergic pathways
The noradrenergic pathways originate in the midbrain locus coeruleus and lateral tegmental area. Noradrenergic pathways are involved in arousal, attention, memory, vigilance, sleep, and mobilizing the brain and body for action, including the fight-or-flight response.
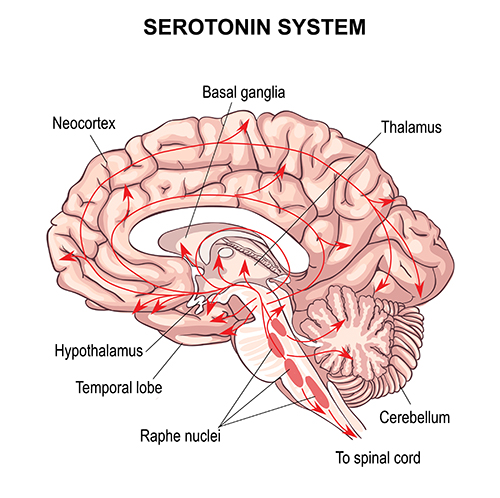
Serotonergic pathways
The serotonergic pathways originate in the brainstem and midbrain raphe nuclei. Serotonergic pathways are involved in appetite, mood, and sleep.
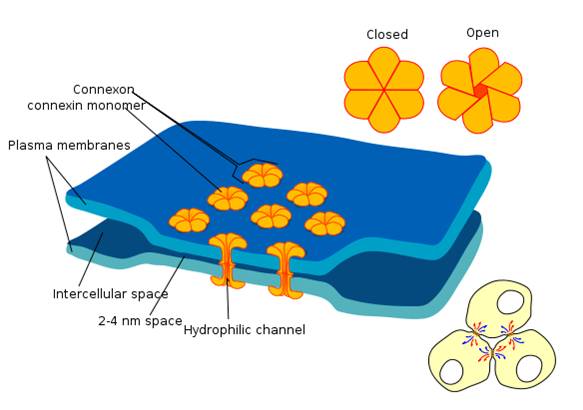
Electrical synapses communicate information across gap junctions
between adjacent membranes using ions. Gap junctions are narrow spaces between two cells bridged by connexons (protein channels) that allow ions near-instantaneous travel. Graphic courtesy of Wikimedia Commons.
Electrical synapses are symmetrical. Ions flow across a 3-nm gap junction into the more negatively charged neuron
as long as the gap junction remains open. This means that whether neurons are presynaptic or postsynaptic
depends on their respective charges. When two neurons are electrically coupled, an action potential in one induces a postsynaptic potential (PSP) in the paired neuron.
Transmission across electrical synapses is instantaneous, compared with the 10-ms
or longer delay in chemical synapses. The rapid information transmission that characterizes electrical synapses
enables large circuits of neurons to synchronize their activity and simultaneously fire.
Gap junctions may be a preliminary step toward developing chemical synapses between these neurons, eventually replacing their electrical synapses. Prenatally and postnatally, gap junctions enable nearby neurons to coordinate their development by sharing electrical and chemical communications (Bear, Connors, & Paradiso, 2020; Breedlove & Watson, 2020).
Old school view: synapses are either
electrical or chemical.
New school view: synapses can be both electrical and chemical
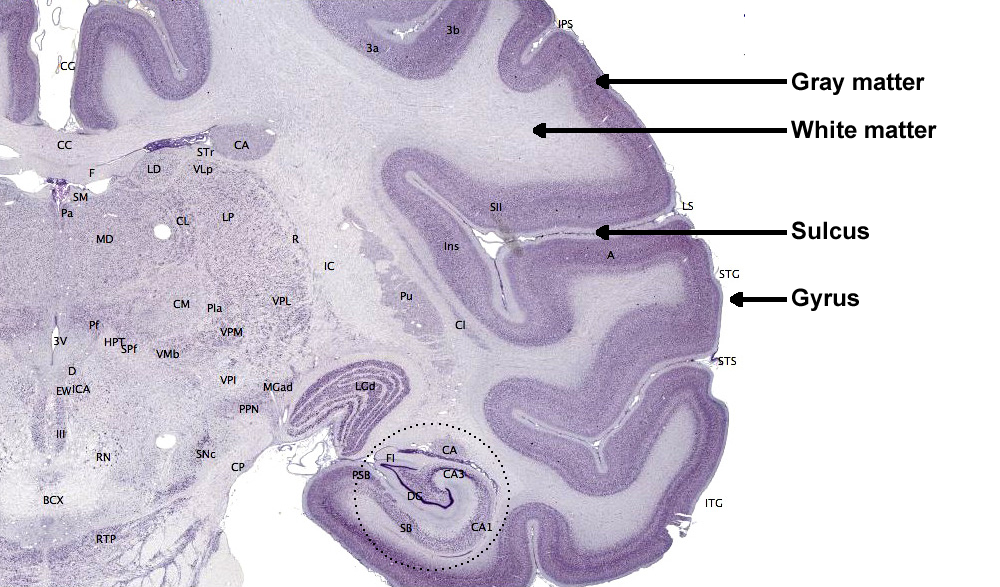
The cerebral cortex comprises neuronal cell bodies, glial cells, and blood vessels. Beneath the neocortex lies myelinated nerves (white matter), unmyelinated fibers, and glial cells.
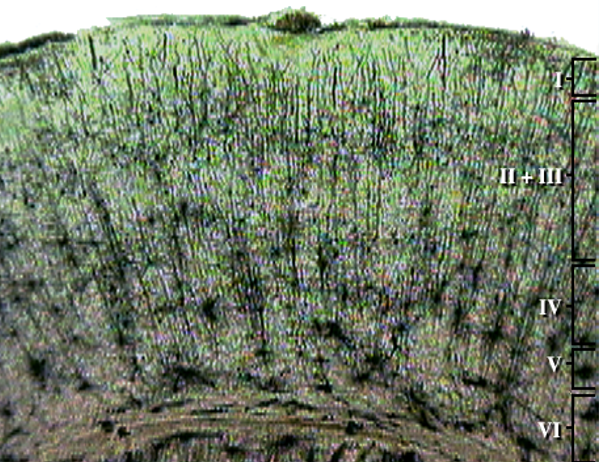
The cerebral cortex covers the cerebral hemispheres and consists of gray and white matter. Gray (or grey) matter,
which looks grayish brown, comprises cell bodies. White matter gains its opaque white color from myelinated axons. The cerebral cortex of a macaque brain is shown below,
courtesy of Wikipedia. Note that staining imparts a darker shade to gray matter.
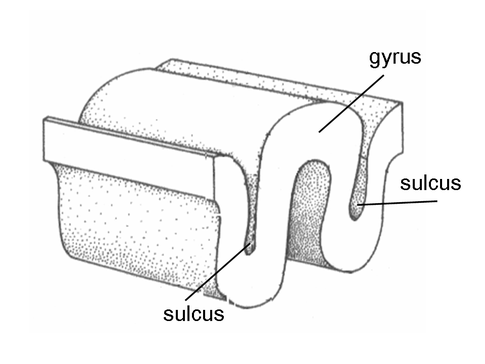
The convolutions of the cerebral cortex contain two-thirds of its surface area and maximize the volume of cortical
tissue housed within the skull. Cerebral cortical convolutions include sulci, which are shallow grooves in the surface of the cerebral hemisphere (central sulcus),
fissures, which are deep grooves (lateral fissure), and gyri, which are ridges of cortex demarcated by sulci or fissures (precentral gyrus)
(Carlson & Birkett, 2021).
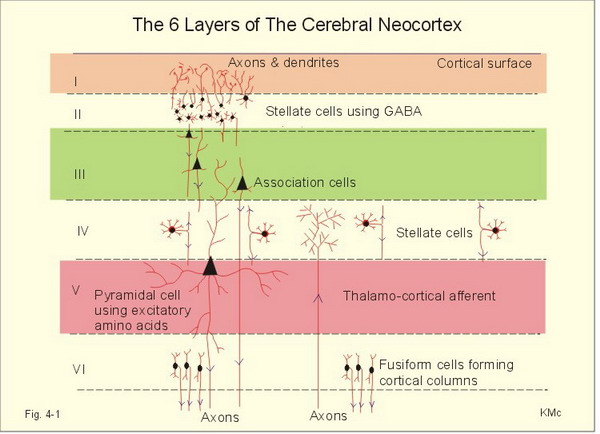
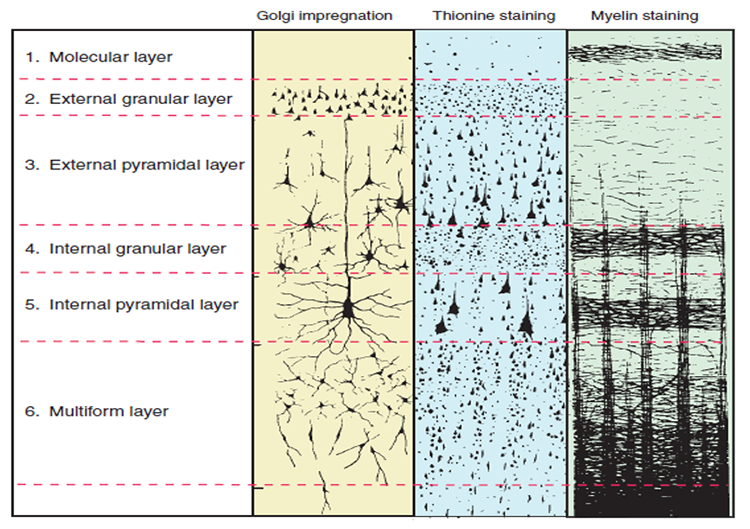
There are two main types of cortex: neocortex and allocortex.
The neocortex or isocortex consists of six layers 3 mm thick with a surface area of about 2360 cm2
with white matter underneath. Layers I-III receive corticocortical afferent fibers
that connect the left and right hemispheres. Layer III is the main source of
corticocortical efferent fibers. Layer IV is the primary destination of thalamocortical
afferents and intra-hemispheric corticocortical afferents. Layer V is the primary
origin of efferent fibers that target subcortical structures that have motor functions. Layer VI
projects corticothalamic efferent fibers to the thalamus, which together with the thalamocortical afferents,
creates a dynamic and reciprocal relationship between these two structures (Creutzfeldt, 1995). Diagram courtesy of Wikimedia Commons.
Allocortex, which means other cortex, usually has between three or four layers, compared with the neocortex's six layers. The allocortex has less volume than the neocortex and comprises the olfactory system and hippocampus.
A transitional region between the neocortex and allocortex is called the paralimbic cortex.
For a basic overview of the cortex, watch the Khan Academy video Cerebral Cortex.
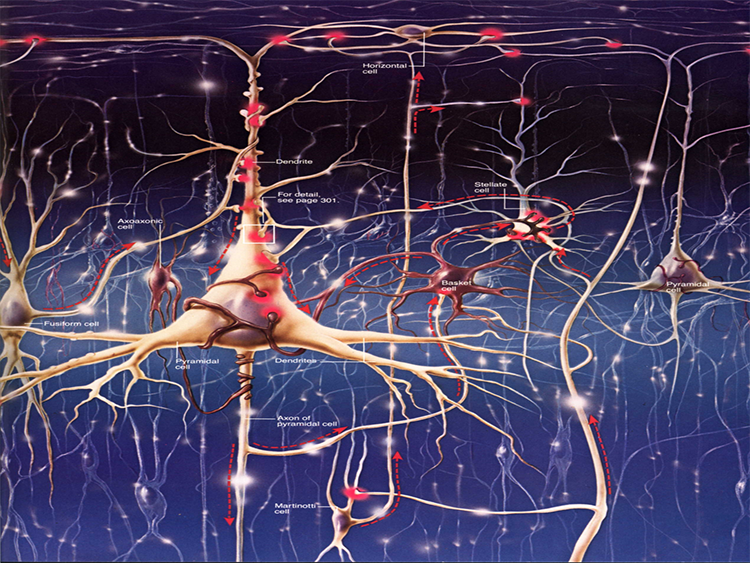
Neurons in the Cortex
We can classify cerebral cortical neurons as whether their dendrites display spines or not. Spiny neurons, which have either pyramidal or stellate (star-like)-shaped cell bodies, are usually excitatory. While all pyramidal cells are spiny neurons, stellate cells can be spiny or aspinous (Bear, Connors, & Paradiso, 2020).
The graphic below depicts dendritic spines.
There are many types of aspinous (smooth) neurons, which are believed to be inhibitory.
Local activity is a composite of local and network influences. Network communication systems and local cortical functions show different characteristics across the cortex and produce unique and specific EEG patterns in other regions.
With the EEG, we can follow the progression from stimulus to behavior response. This allows us to determine the correct function at each step and identify causal factors in dysfunctional outcomes or responses.
Source of the Scalp EEG
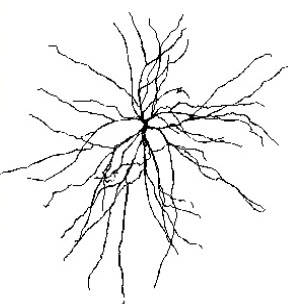
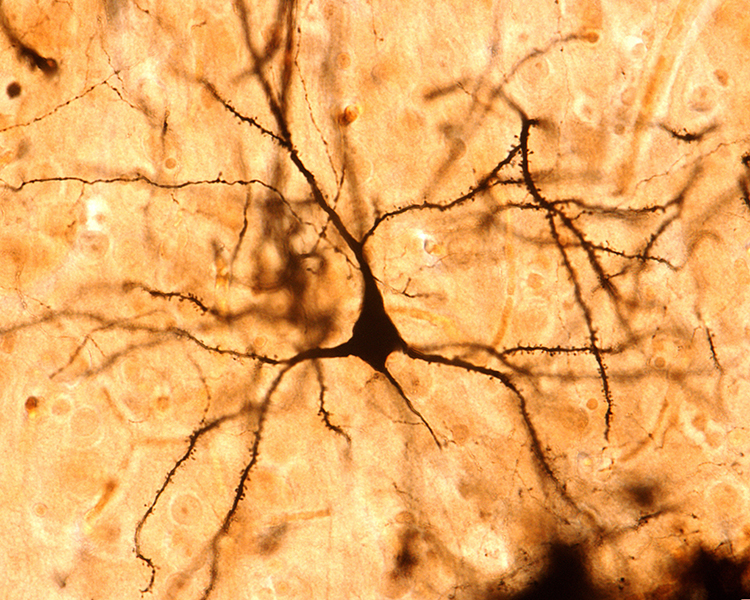
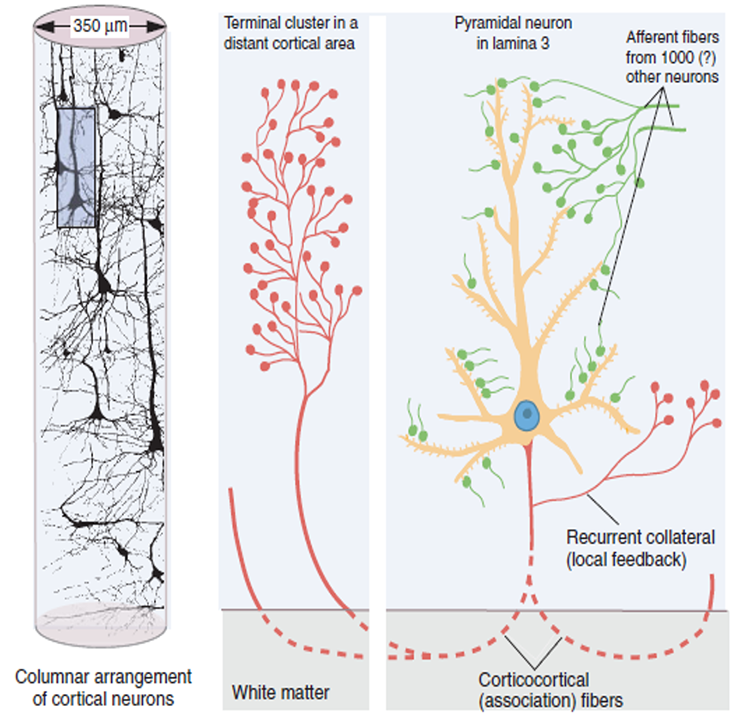
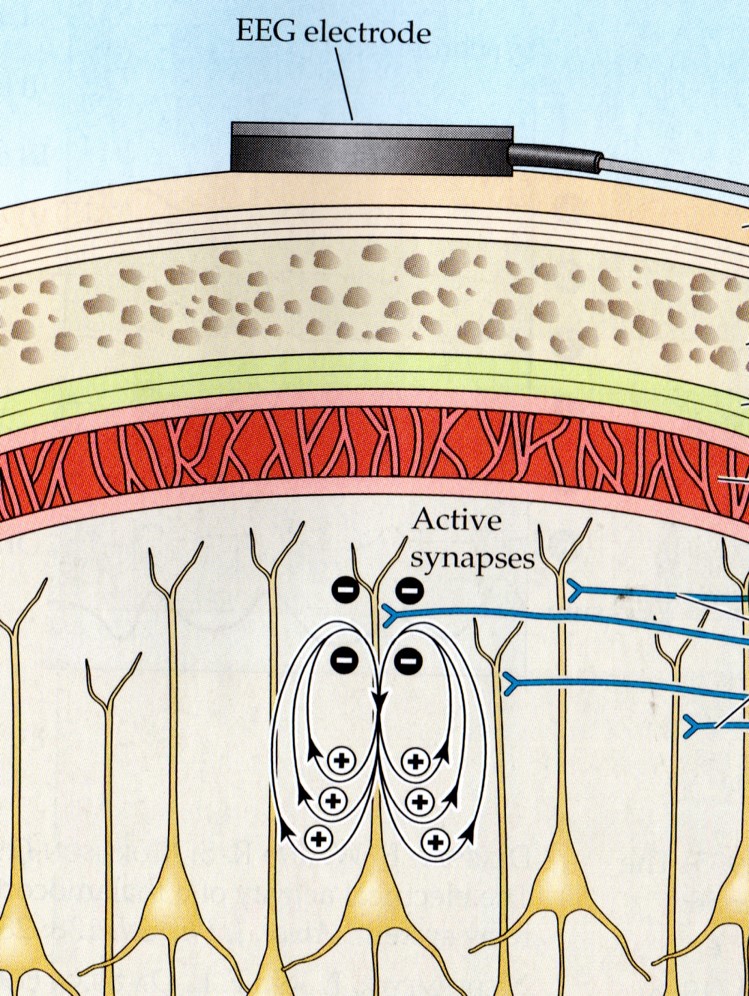
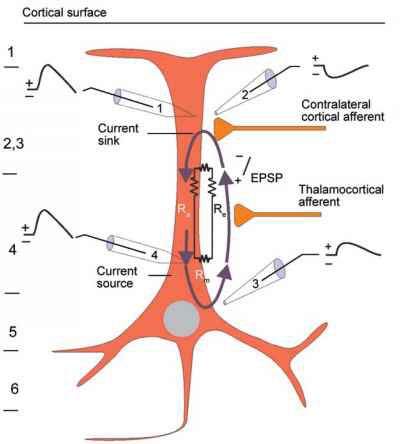
The scalp EEG results from the summation of large areas of gray matter activity. Areas are polarized synchronously due to the input of oscillatory or transient evoked activity. These areas comprise thousands of cortical columns containing large pyramidal cells aligned perpendicularly to the cortical surface.
Pyramidal neurons are found in all cortical layers, except layer 1, and represent the primary type of output neuron in the cerebral cortex.
The scalp EEG results from the summation of EPSPs and IPSPs in thousands of cortical columns containing large pyramidal cells perpendicular to the cortical surface. The columns are synchronously polarized (made more negative) and depolarized (made less negative) due to the input of oscillatory or transient evoked activity.
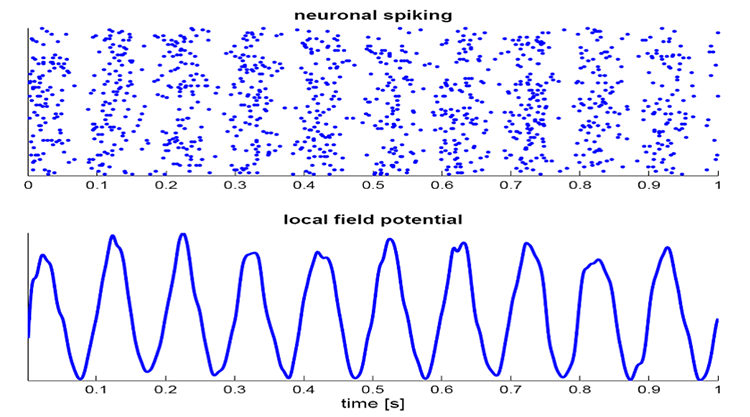
Local Field Potentials
The local field potential (LFP) is the aggregate effect of the firing of the interconnected pyramidal neurons within the cortical columns, plus additional mechanisms like glial cell modulation of the cortical electrical gradient.
Caption from Wikipedia's article on Neural Oscillation. Simulation of neural oscillations at 10 Hz. The upper panel shows spiking of individual neurons (with each dot representing an individual action potential within the population of neurons). On the lower panel, the local field potential reflects their summed activity. This figure illustrates how synchronized patterns of action potentials may result in macroscopic oscillations that can be measured outside the scalp.
Do not confuse the "spiking" of individual neurons with epileptogenic spikes in the scalp EEG.
Scalp Electrical Potentials
Scalp electrical potentials represent the sum of all available electrical fields. Fields of opposite polarity (+/-) cancel each other out so that scalp potentials are greater when large aggregates of neurons polarize and depolarize synchronously. The scalp EEG represents a weighted sum of all active currents with the brain that generate open fields, including non-cortical sources.
Action potentials reflect neuronal output. They are seen in extracellular recordings as fast (~300 Hz)
activity that exceeds 90 mV lasting less than 2 ms. Action potentials play a minor role in scalp surface EEG. They fall below 60 V outside of a 50-μm (0.050-mm) radius. Scalp electrodes are several centimeters from cortical neurons and are generally aligned away from the scalp. Therefore, action potentials are unlikely to contribute significant voltages to the scalp EEG.
Local Field Potentials Regulate Neuron Excitability and Firing
Neurons are most likely to fire during the depolarizing phase of the local field potential. Neurons are more excitable when they are "in phase" with respect to the local field potential (LFP) and are inhibited when they are out of phase with the LFP. Thus, at any instant of time, the amplitude and frequency of the EEG are regulated by the LFP, which in turn, is influenced by oscillatory mechanisms such as slow cortical potentials.
The EEG is a moment-to-moment measure of the excitability of action potential firing, like gates opening and closing on the half cycle.
The synchronous activity of large pyramidal neurons networked in cortical columns creates the EEG.
The Composition of the EEG
The EEG is composed of electrical potentials, varying in two dimensions, frequency and amplitude.
Sources of IPSP and EPSP Inputs
Many sources contribute input that results in IPSP and EPSP activity within cortical neurons. These sources primarily contribute influences such as oscillatory generator input or ascending event-related evoked input.
The
eyes open again at 14’31”, and alpha attenuates (alpha blocking). This shows the posterior dominant rhythm (generally known as "alpha") appearing in the eyes-closed condition when visual sensory input is stopped. The attenuation or blocking of this rhythm as sensory input returns in the eyes-open condition.
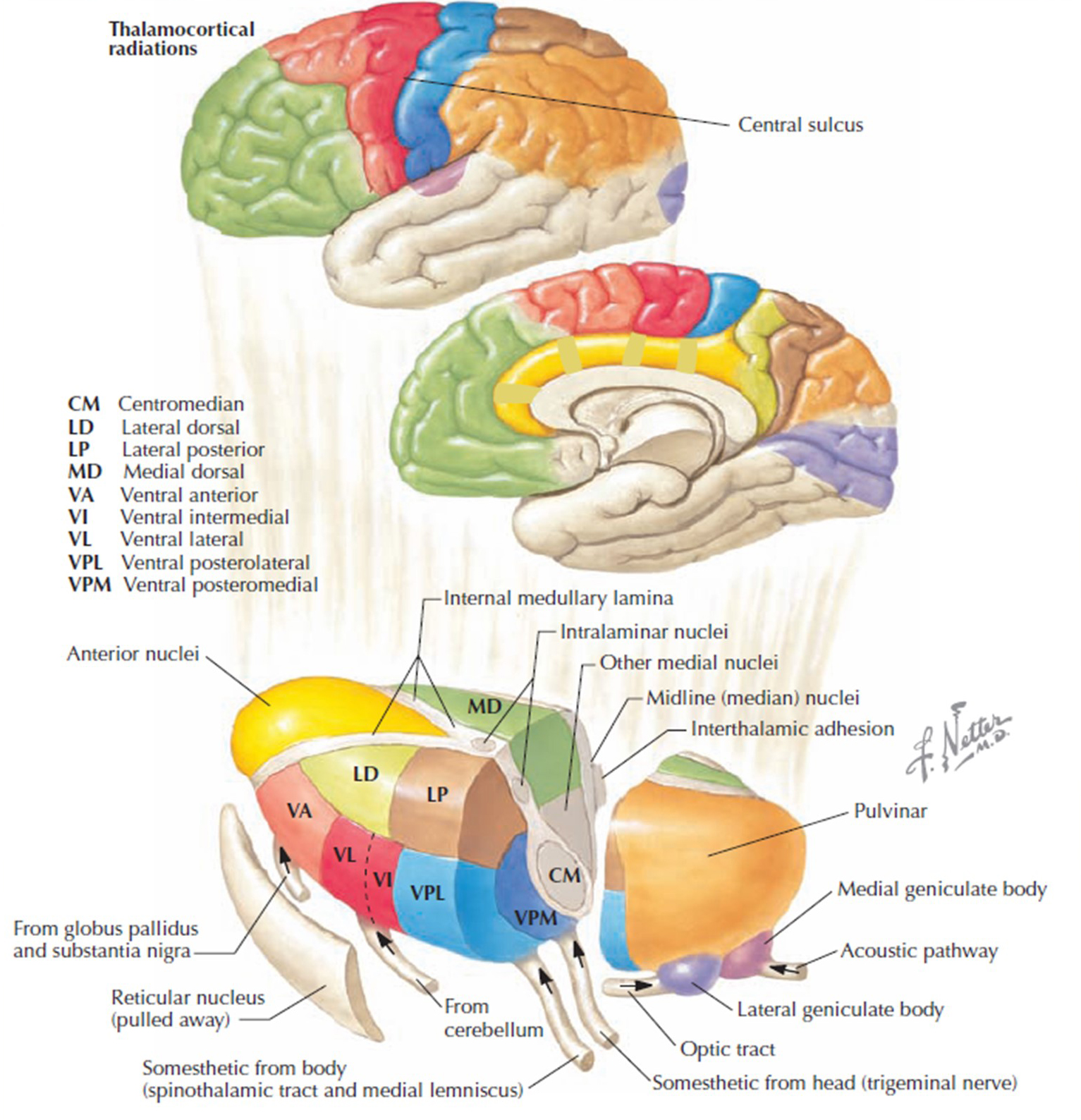
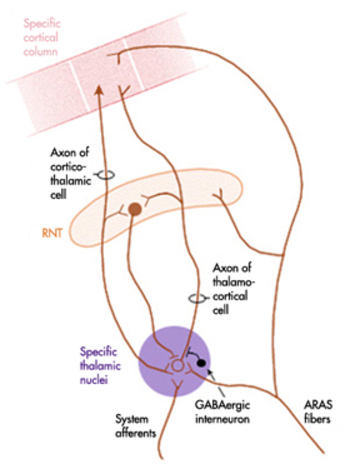
The diagram below, which shows bidirectional connections between the thalamus and cortex, was modified from the original on www.lib.mcg.edu.
Caption by W. D. Jackson, PhD, and S. D. Stoney, PhD (2006): Thalamocortical cells are subject to excitatory drive from their system afferents, from monosynaptic corticothalamic fibers, and from the brainstem reticular formation (ascending reticular activating system, ARAS). They receive inhibitory drive from local interneurons and neurons in the reticular nucleus of the thalamus (RNT). Note that the RNT neurons are excited by activity in thalamocortical cells and corticothalamic cells. The connections are precisely organized. For example, each column in a primary cortical area sends corticothalamic fibers back to the same part of its specific thalamic nucleus that sends its thalamocortical fibers to that cortical column. The corticothalamic fibers also synapse on the RNT cells receiving input from that part of the thalamic nucleus. Each cortical receiving area is said to be "reciprocally connected" with its specific thalamic nucleus. Like the thalamocortical cells, RNT cells and cortical neurons also receive excitatory drive from the ARAS.
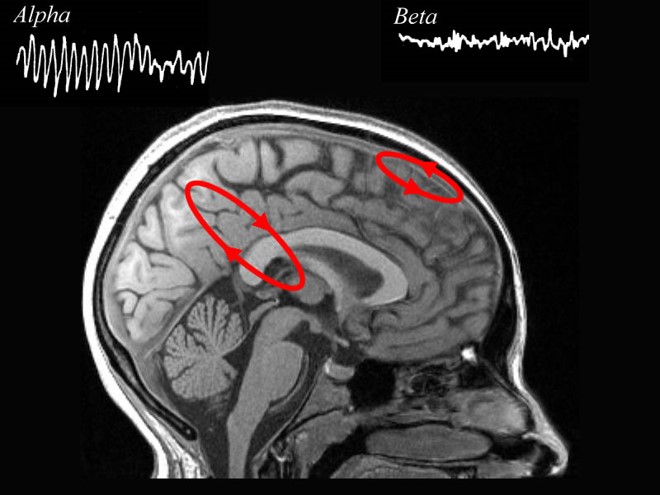
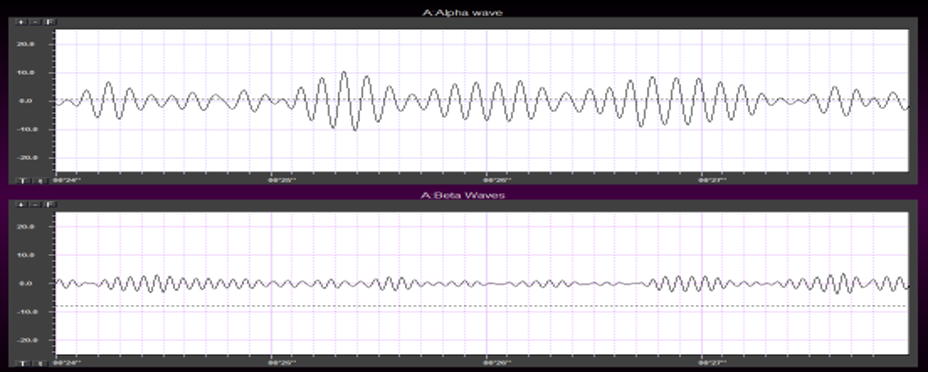
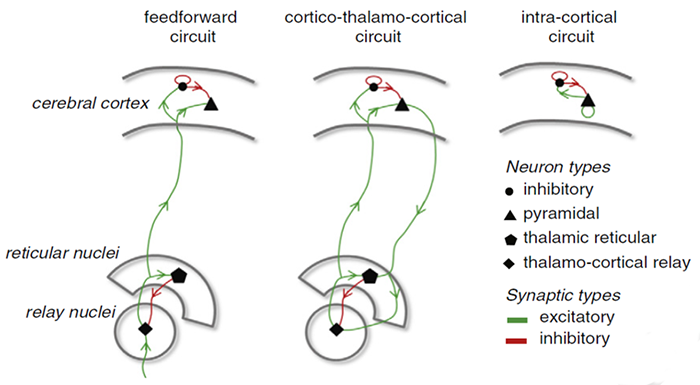
The EEG is generated by thalamocortical (alpha) and cortical-cortical
(beta) sources.
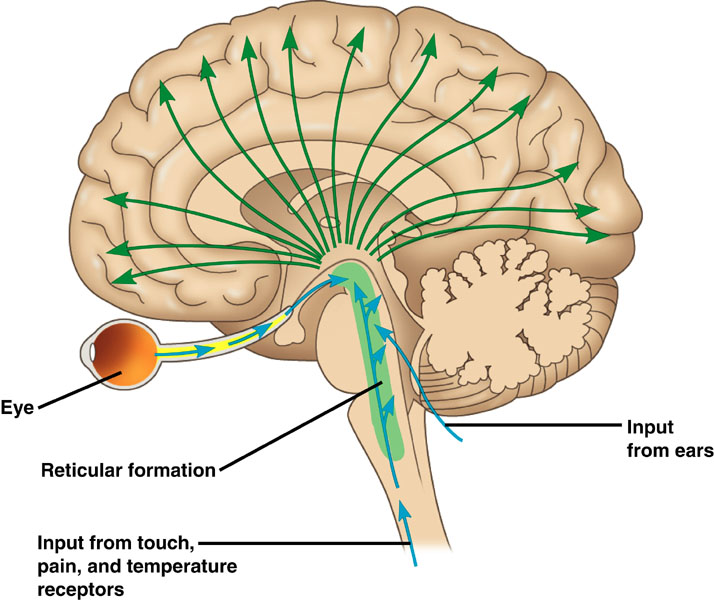
Neurons in the ascending reticular activating system produce event-related potentials in response to diverse stimuli like a flashing light or sound. Event-related potentials (ERPs) are the brain's response to externally applied stimuli, events, or cognitive/motor tasks. They are time-locked measures of brain electrical activity.
Dipole Generators
Large cortical pyramidal neurons organized in macrocolumns are oriented with an apical dendrite projecting toward the scalp and an axon descending in the opposite direction. An "Equivalent Dipole Generator" usually represents the sum of all multipolar current sources. Summed generators are modeled as dipoles to aid the conceptual understanding of the electrical fields involved.
EEG Signals (Brainwaves)
The EEG represents changes in a brain area's electrical activity (potential) compared to a "neutral" site or another brain area. The EEG is displayed as oscillations or voltage fluctuations, which show a "wave" pattern when plotted on a graph.
"These oscillations are generated spontaneously in several areas of the cerebral cortex as neuronal networks transiently form assemblies of synchronously firing cells." Klaus Linkenkaer-Hansen.
Sink, Source, and Dipole
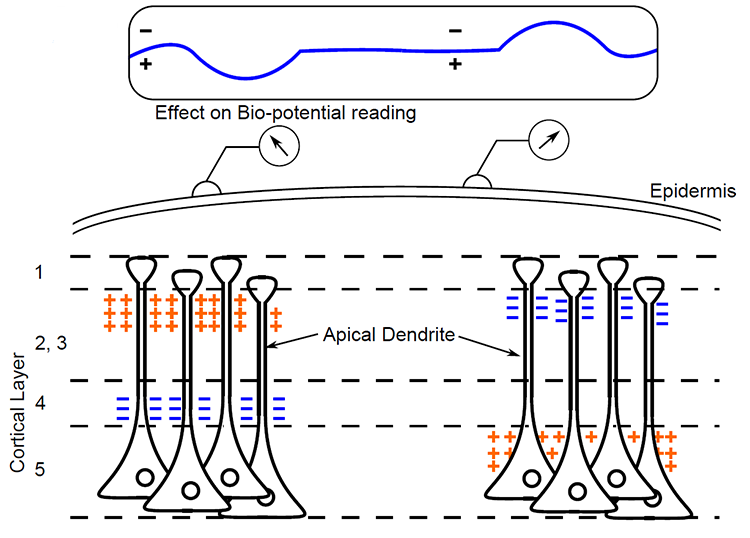
We can describe pyramidal cells in terms of their sink, source, and dipole. A sink (-ve), which may be located at the bottom, middle, or top of the apical dendrite, is where positive ions enter the dendrite. Cation (positive ion) entry gives the extracellular space a negative charge. The source (+ve) is where the current exits the cell. Finally, the dipole is the field created between the sink and source (Thompson & Thompson, 2016).
The graphic below from Euroform Healthcare: Conduction Studies depicts current entering the apical dendrite (sink) of a pyramidal neuron where an afferent neuron has generated an EPSP. The current leaves the neuron (source) from the dendrite or cell body.
The postsynaptic potentials (EPSPs and IPSPs) propagated by the apical dendrites in layers 2 and 3 create an extracellular dipole layer parallel to the cortical surface. The dipole layer's electrical polarity is the opposite of the deeper cortical layers 4 and 5 (Fisch, 1999).
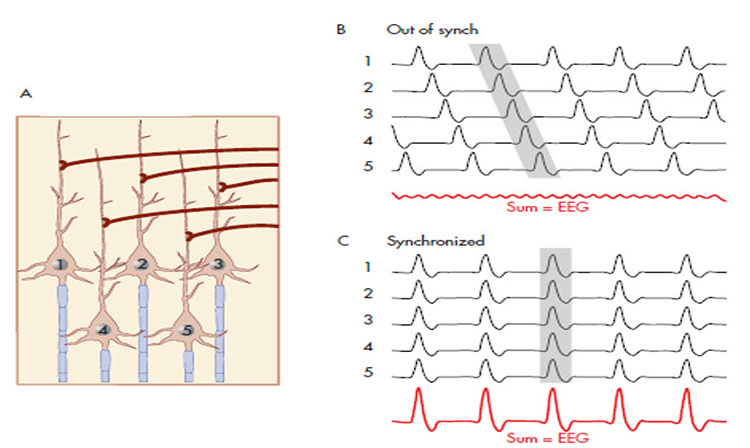
A cortical dipole is created when pyramidal neurons depolarize simultaneously. This phenomenon is called local synchrony. Fewer than 5% of pyramidal neurons can generate more than 90% of the power in the EEG signal because most pyramidal neurons usually fire asynchronously so that their potentials counteract each other. A small fraction of these neurons firing in step can produce visible changes in EEG feedback. This creates the potential for operant conditioning to help clients learn to modify EEG activity through neurofeedback.
Cortical dipoles have three properties: site (depends on source), size (oscillation frequency and voltage), and relative position with respect to sulci and gyri (Collura, 2014).
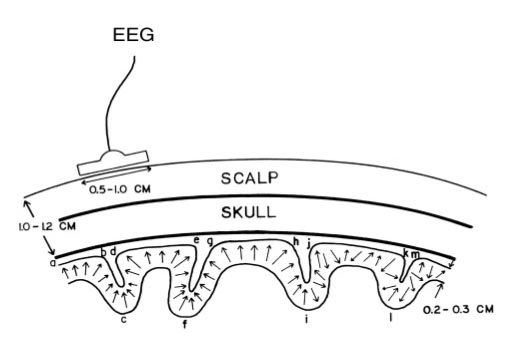
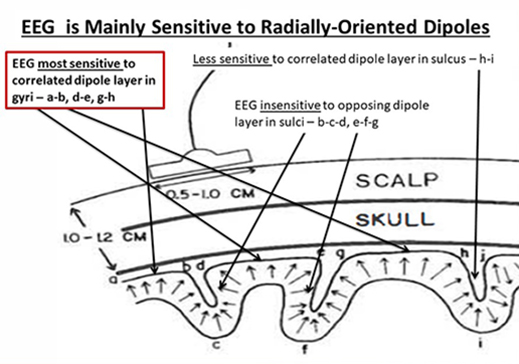
The EEG is Mainly Sensitive to Radially Oriented Dipoles
Evolution has convoluted the human brain to increase its computing power without enlarging the skull. This enfolding has created two easily visible anatomical features: gyri and sulci.
Recall that a gyrus is a ridge of the convoluted cerebral cortex, while a sulcus is a valley. The graphic below is courtesy of Wikipedia.com from the article Sulcus (neuroanatomy).
The EEG is composed of electrical potentials that vary along the dimensions of amplitude and frequency.
EEG Amplitude
The "amount" or amplitude and the "pattern" or morphology of any EEG frequency band reflect the number of neurons discharging simultaneously at that frequency. Lower neuron firing rates correspond to lower signal amplitude.
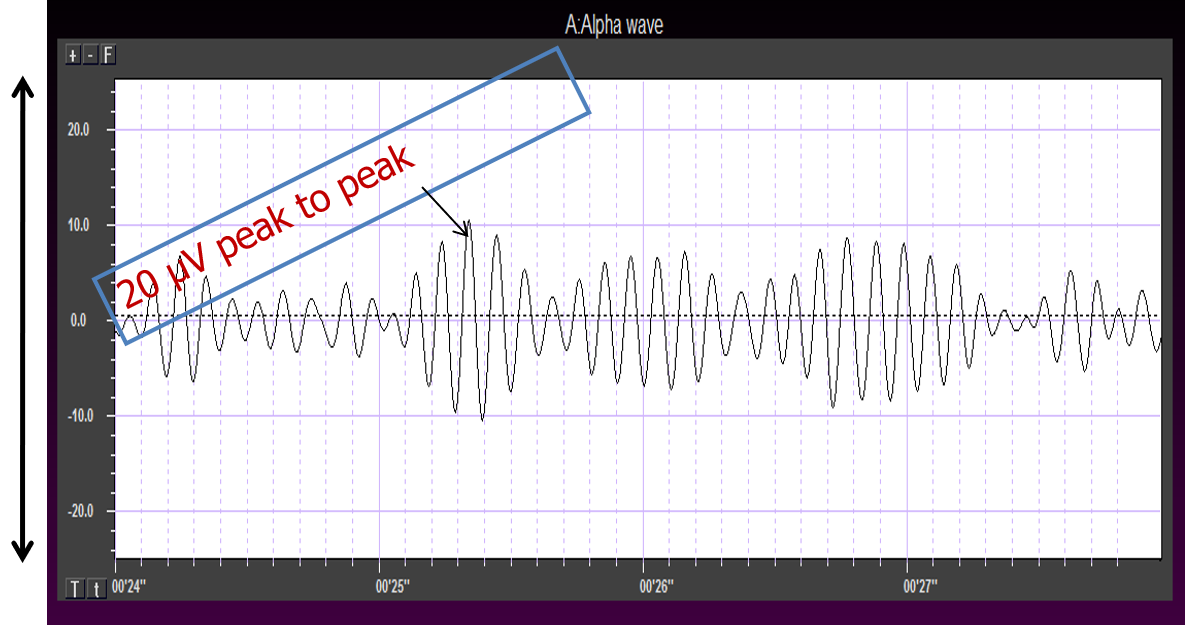
Amplitude measures the amount of energy in the signal and is usually expressed in microvolts.
Greater synchrony in firing among neurons results in higher amplitude, as shown with alpha in the graphic below.
Greater firing synchrony produces larger EEG potentials that can be measured from the scalp surface.
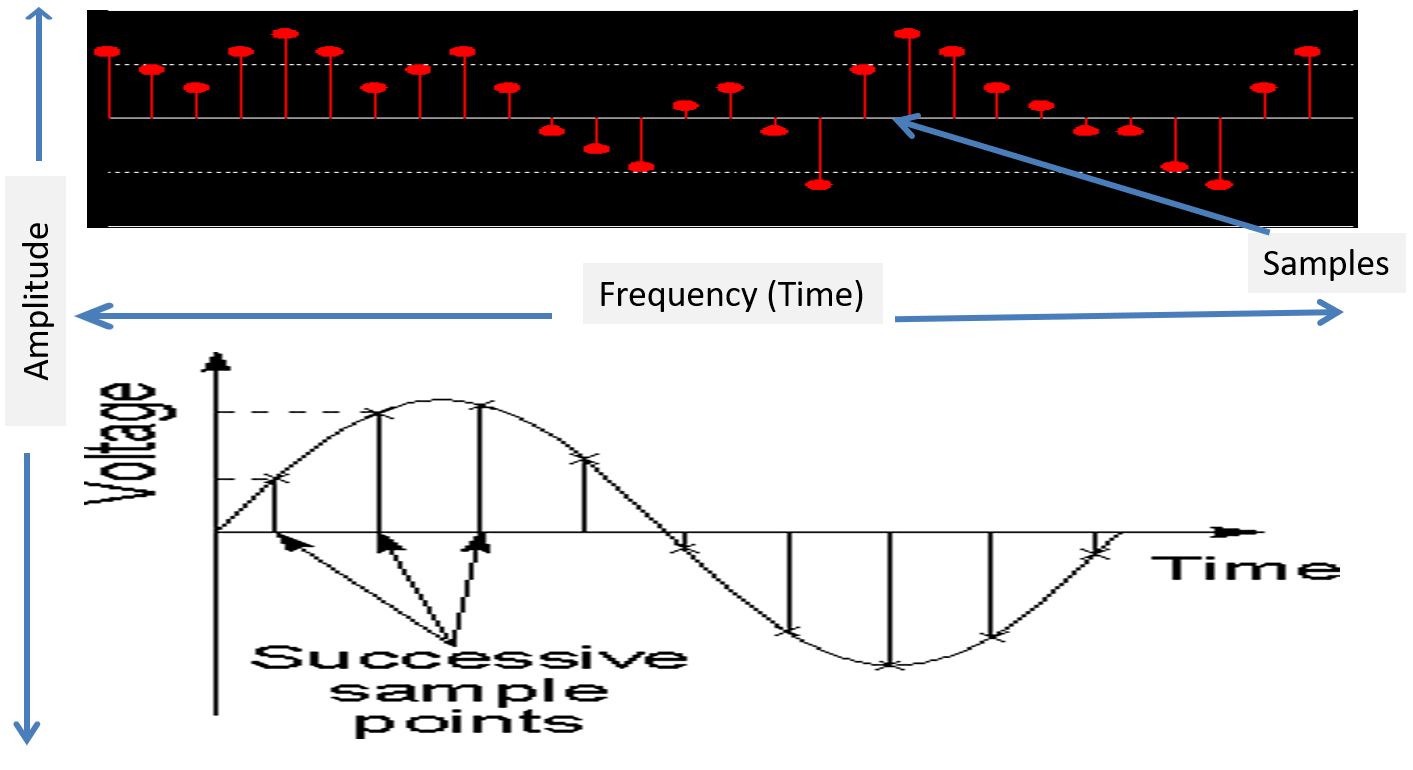
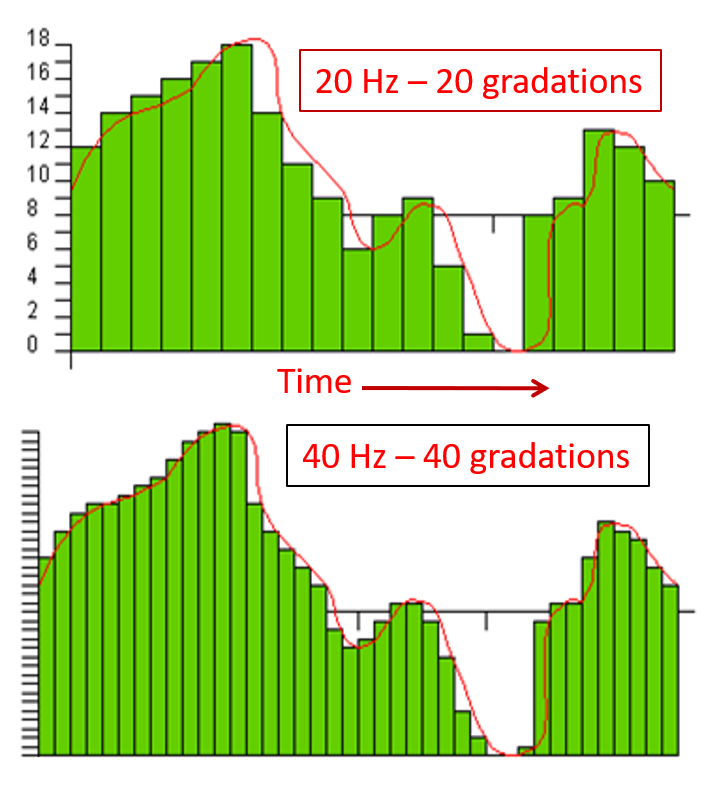
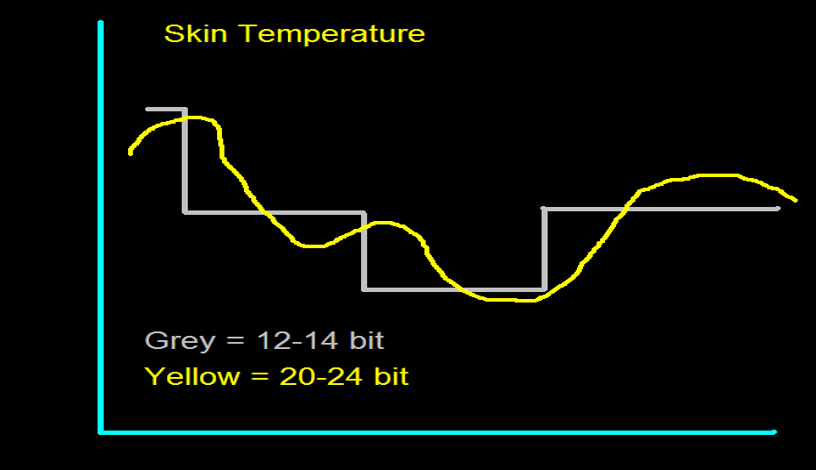
The EEG plots voltage changes over time, which can be displayed on a graph. The sampling rate is the number of measurements per second (Hz). Precision is the number of voltage gradations or steps.
The analog-to-digital (A/D) converters that transform voltages into numerical values vary in precision: more bits correspond to greater accuracy. The graphic below shows precision differences using 12-14-bit (grey) and 20-24-bit A/D converters.
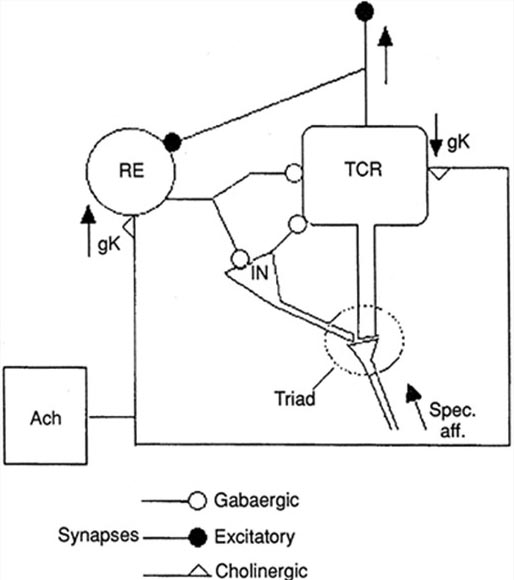
The generation of oscillatory activity, sometimes called spindle behavior, is likely due to the interaction between thalamocortical relay neurons (TCR), reticular nucleus neurons (RE), and interneurons. These interactions are mediated by diverse neurotransmitters, including acetylcholine and GABA.
The various spindle frequencies, which have often been interpreted as reflecting different types of oscillations, merely depend on various durations of the hyperpolarizations (negative shifts) in thalamic-cortical relay neurons. Long duration hyperpolarizations, as during ... deeply EEG-synchronized states, are associated with 7 Hz or even lower-frequency spindles, while relatively short hyperpolarizations result in ... higher frequencies (14 Hz) (Steriade, 2005).
The Purpose of Oscillatory Activity
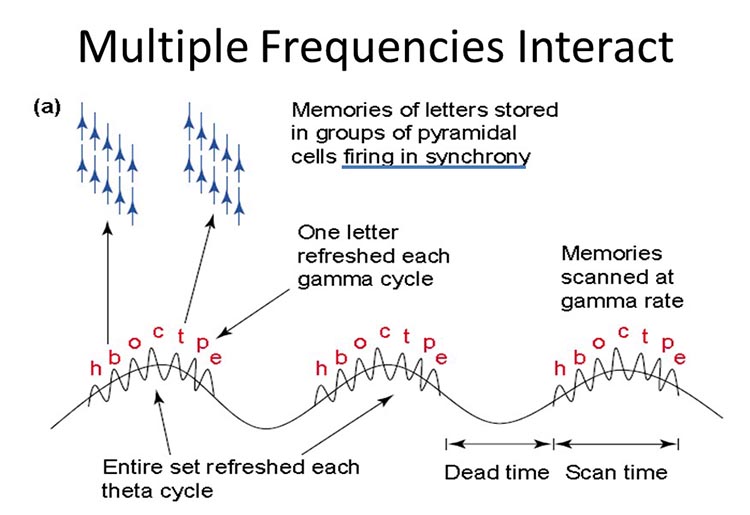
A single neuron can influence multiple postsynaptic targets located between 0.5 and 5 mm away with conduction periods of between 1 and 10 ms. This time difference becomes progressively more pronounced when more complex events involve progressively larger assemblies of neurons. It may take hundreds of thousands of neurons, stimulating multiple postsynaptic neurons, for the desired outcome to occur. When these many neurons are involved, it becomes increasingly clear that there is a need for organization and structure to manage this diverse activity.
Timing is everything since action potentials arrive from a large number of sources. The nervous system must correctly register arrival times to recognize a face, recall a name, or remember personal history and context.
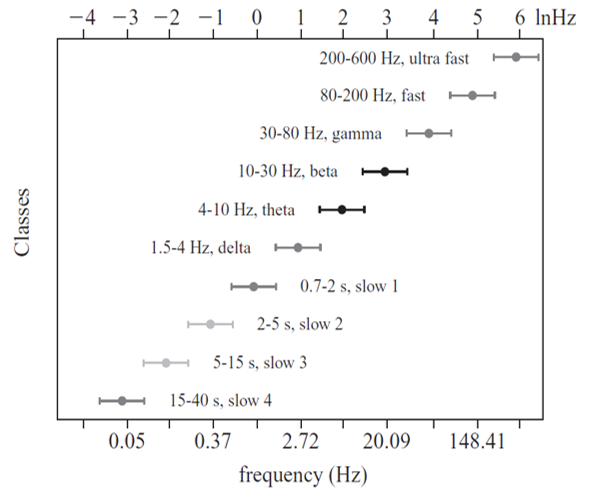
"Oscillatory classes in the cerebral cortex show a linear progression of the frequency classes on the log scale. In each class, the frequency ranges ('bandwidth') overlap with those of the neighboring classes, so that frequency coverage is more than four orders of magnitude" (Buzaki, 2006).
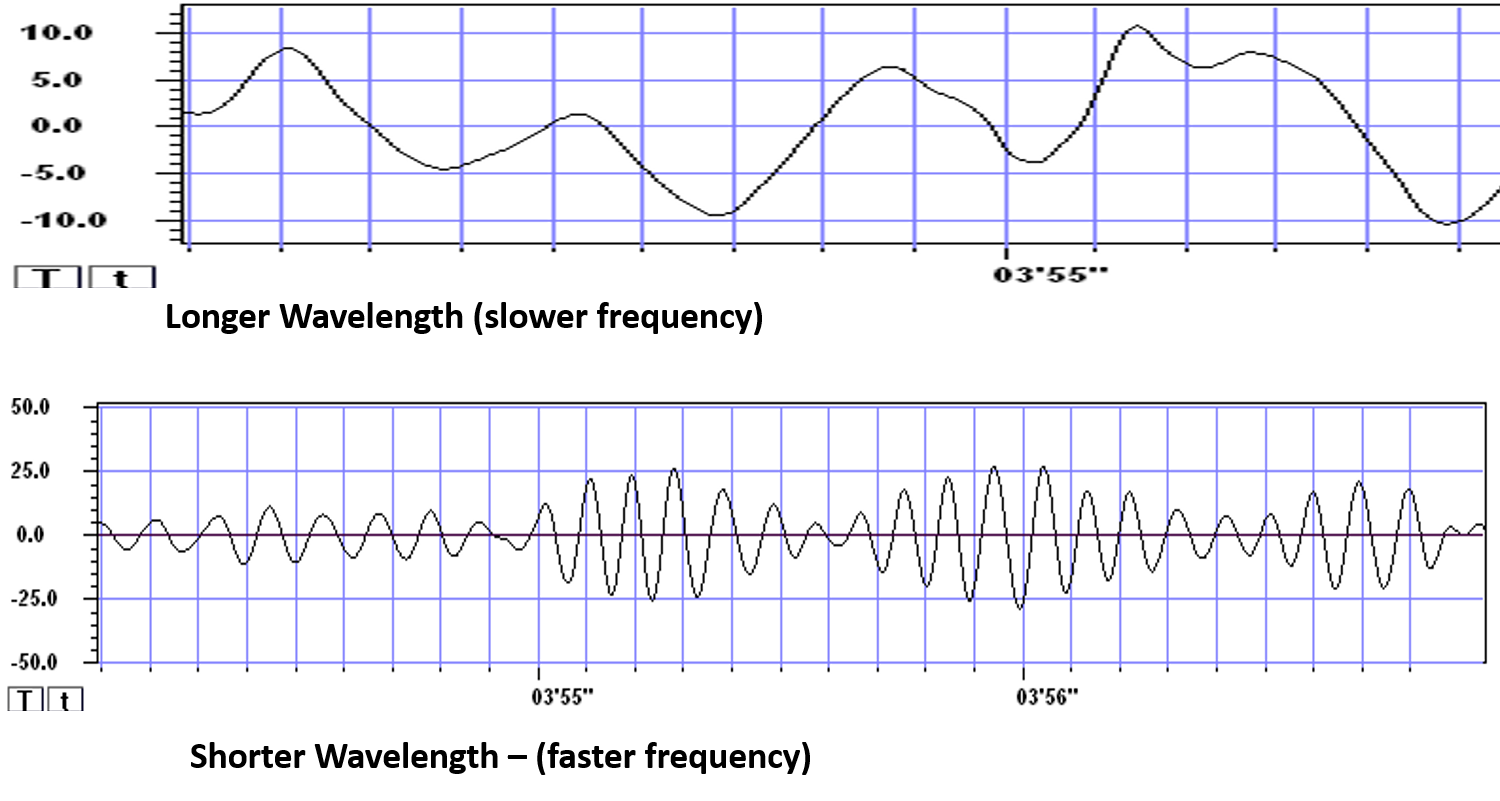
Frequency Determines Complexity
The wavelength or frequency of the EEG band determines how long the processing window will remain open and, therefore, the size of the neuronal pool involved. Because of the distances involved, longer wavelengths (slower frequencies) allow larger groups of more distant neurons to be assembled and coordinated. Different frequencies organize different types of connections and different levels of computational complexity.
Short time windows of fast oscillators facilitate local integration, primarily because of the limitations of axon conduction delays. Fast oscillations favor local decisions. Slow oscillators can involve many neurons in large and/or distant brain areas. Slow oscillations favor complex, global decisions.
Complexity Versus Frequency
Complex tasks involving sensory integration and decision-making were associated with 4-7 Hz synchronization. Intermediate tasks such as identifying spoken and written words and pictures increased 13-18 Hz beta activity. Simpler, more localized tasks, such as the visual processing of grid displays, were associated with faster-frequency activity (24-32 Hz) (Sarnthein et al., 1998; Von Stein et al., 1999).
Traveling Waves Help Coordinate Widespread Brain Networks
Zhang et al. (2018) proposed that traveling waves between 2 to 15 Hz, moving at 0.25-0.75 meters per second across the cortex, mediate large-scale coordination of brain networks and support connectivity.
Summary of EEG Oscillations
When the CNS processes incoming content, separate areas detect features of salient content, including visual, auditory, tactile, kinesthetic, and olfactory information. The CNS shares, integrates, compares current with previous content, analyzes, and makes decisions regarding memory and responses. Interacting networks linked by electrical and chemical signals perform this work. We record the electrical potentials generated by this complex and dynamic network activity as the EEG.
Sensory evoked potentials are a subset of event-related potentials (ERPs)
Event-related potentials (ERPs) represent the brain's responses to external stimuli, events, or cognitive/motor tasks. ERPs can be detected throughout the cortex.
Investigators monitor ERPs by placing electrodes at areas like the midline (Fz, Cz, and Pz). A computer analyzes a subject's EEG responses to the same stimulus or task over many trials to subtract random EEG activity. ERPs always have the same waveform morphology. Their negative and positive peaks occur at regular intervals following the stimulus.
Sensory evoked potentials are a subset of ERPs
elicited by external
sensory stimuli (auditory, olfactory, somatosensory, and visual). They have a negative peak at
80-90 ms and a positive peak at about 170 ms following stimulus onset. The orienting
response ("What is it?") is
a sensory ERP. The N1-P2 complex in
the auditory cortex of the temporal cortex reveals whether an
uncommunicative person can hear a stimulus.
Motor ERPs are detected over the
primary motor cortex (precentral gyrus) during movement, and their
amplitude is proportional to the force and rate of skeletal muscle
contraction (Thompson & Thompson, 2016).
Slow cortical potentials modulate the excitability of associated neurons
Slow cortical potentials (SCPs) are
gradual changes in the membrane potentials of cortical dendrites that
last from 300 ms to several seconds. These potentials include the
contingent negative variation (CNV), readiness potential,
movement-related potentials (MRPs), and P300 and N400 potentials, and exclude event-related potentials (ERPS)
(Andreassi, 2007).
SCPs modulate the firing rate of cortical pyramidal neurons by exciting
or inhibiting their apical dendrites. They group the classical EEG rhythms using these synchronizing mechanisms
(Steriade, 2005).
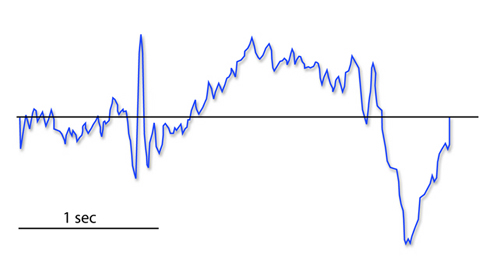
The contingent negative variation (CNV) is a steady, negative shift in
potential (15 µV in young adults) detected at the vertex. This
slow cortical potential may reflect expectancy, motivation, intention to
act, or attention. The CNV appears 200-400 ms after a warning signal
(S1), peaks within 400-900 ms, and sharply declines after a second
stimulus that requires the performance of a response (S2). John Balven adapted the graphic below from Stern, Ray,
and Quigley (2001).
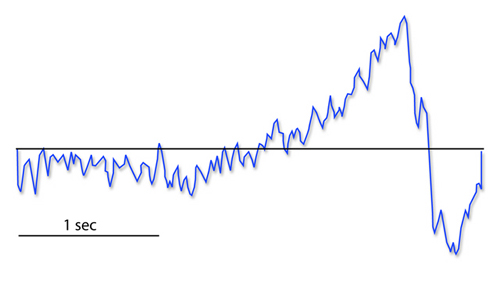
The readiness potential is a slow-rising, negative potential (10-15
µV) detected at the vertex before voluntary and spontaneous
movement. This slow cortical potential precedes voluntary movement by 0.5
to 1 second and peaks when the subject responds. This potential is
separate from the CNV. John Balven adapted the graphic below from Stern, Ray, and Quigley (2001).
Movement-related potentials (MRPs) occur at 1 second as subjects prepare for
unilateral voluntary movements.
MRPs are distributed bilaterally with maximum amplitude at Cz. The
supplementary motor area and primary motor and somatosensory cortices
generate these potentials (Babiloni et al., 2002).
P300 and N400 ERPs are classified as long-latency potentials due to their
extended latencies
following stimulus onset.
The P300 potential is an event-related potential (ERP) with a 300-900-ms
latency.
The largest amplitude positive peaks are located over the parietal lobe. Researchers
elicit the P300 potential by exposing subjects to an odd-ball stimulus, a
meaningful
stimulus that is different from others in a series (a colored playing card
presented in a series of monochrome cards). The P300
potential may reflect an event’s subjective probability, meaning, and
transmission of information. Research shows this is separate from
the contingent negative variation (CNV) (Stern, Ray, & Quigley, 2001).
Shorter P300 latencies may reflect better allocation of attention, and
researchers have measured longer P300 latencies in ADD than non-ADD
samples. Experimental subjects show longer latencies when lying than when
telling the truth (Farwell & Donchin, 1991; Thompson & Thompson, 2016).
The N400 potential is an
event-related potential (ERP) elicited when we encounter semantic
violations like ending a sentence with a semantically incongruent word
("The handsome prince married the beautiful fish"),
or when the second word of a pair is unrelated to the first (BATTLE/GIRL). Warren and
McIlvane (1998) speculate that the N400 potential is evoked
whenever a general conceptual system that produces category judgments
encounters a mismatch that violates equivalence relations. Halgren and
colleagues (2002) consider it an index of the difficulty of semantic
processing.
A Deep Dive Into SCPs
In 1875, Richard Caton identified what may have been the first evidence of SCPs in an article in the British Medical Journal titled "The Electric Currents of the Brain."
He stated, "The cortex's Direct Current baseline waxes negative whenever it is more active. Gradients of 150-200 μV/mm are noted." He later noted, "when any part of the gray matter is in a state of functional activity, its electric current usually exhibits negative variation." Some later researchers suggested that this signaled the discovery of the "steady potential" or the DC potential of the brain. However, others have noted the possibility of equipment-based artifacts in his recordings (Niedermeyer, 1999).
From the late 1800s through the early 1900s, research into brain electrical activity turned toward observations of electrical stimulation and spontaneous electrical activity in animal studies. As technology improved, the ability of researchers to identify EEG rhythms also improved. Hans Berger is famous for his description of alpha-blocking with cognitive activity, made possible partly because of his use of more sensitive equipment (Niedermeyer, 1999).
Subsequent research into the electrical characteristics of the human brain became primarily focused on phasic phenomena from AC-coupled recordings. This trend continues today with the current practice of EEG biofeedback or neurofeedback, focusing primarily on training AC frequencies, generally in the range of 1 to 60 Hz.
The study of SCPs continued in physiology and animal research. Only recently has there been increased interest in observing SCPl values in the human EEG and correlating them with cognitive activity, sensory processing, and motor activity. SCPs are distinguished from short-latency, event-related potentials (ERP) up to 500 ms. SCPs reflect cortical processes that require more than one second to complete and are associated with more global, task-related activities. Such changes occur in task-specific areas of the cortex and can be displayed using topographic maps. Areas of activation show surface negative potential changes (Altenmuller & Gerloff, 1999).
Operant conditioning of SCP changes is an even more recent study area. One reason for increased interest in SCP training is the excellent work done by Birbaumer and colleagues (1999) at the University of Tubingen in Germany, demonstrating that SCPs can be operantly conditioned with positive outcomes for a variety of disorders. The recent availability of DC-coupled amplifiers for EEG recording has also contributed to this interest (Altenmuller & Gerloff, 1999).
According to Niedermeyer (1999), "DC" can mean several things. DC means direct current, which is a current without oscillations. From an electrophysiological perspective, "DC shifts" are ultra-slow potentials below the typical EEG in the oscillation frequency and are generally around 0.1-0.2 cycles per second. However, they may extend up to 1 cycle per second. So SCPs are not true direct current, though their oscillations are so slow that they are "DC-like" phenomena.
DC also refers to "direct coupling" (Niedermeyer, 1999) and describes a type of amplifier that does not use capacitors between the amplification stages and uses an infinite time constant to provide for optimal DC recording. Until recently, this has been quite difficult to achieve for EEG recording. Most conventional EEG amplifiers use capacitors in the input stage, which reject DC voltages and create a finite time constant that interferes with access to DC phenomena.
An approximation of DC information can be obtained from an alternating current amplifier by using a rectifier or extending the time constant to approximately 10 seconds (Kotchoubey et al., 1999). A thorough discussion of amplifier characteristics is beyond the scope of this article. Several excellent chapters relating to this subject can be found in Niedermeyer and Lopes da Silva (1999).
The source and nature of slow cortical potentials are in dispute. The prevailing theory holds that negative SCPs result from synchronous postsynaptic potentials in the apical dendrites of cortical pyramidal cells. Others hold that SCPs are supported and produced by glial cells within the cortex. It appears that pyramidal neurons may be the source of these potentials and that the glial system is the "sink" in electrical terms (Strehl, 2005, personal communication).
Increased neuronal activity is associated with an increased outflow of potassium ions leading to increased extracellular potassium concentrations. Glial cells depolarize when extracellular potassium concentrations increase, resulting in intracellular and extracellular current flows similar to typical neuronal synaptic transmissions (Speckmann & Elger, 1999). Since glial cells are widely interconnected and have extensive processes, it appears likely that the glial system is responsible for the potential changes that produce SCP values recorded from the scalp in response to neuronal activity.
Despite some discussion regarding the source of SCP activity, it is clear that scalp SCPs represent the cortex's excitability potential. SCP negativity is associated with increased cortical excitability. High cortical negativity has been correlated with a greater likelihood of seizures (Speckmann et al., 1984) and migraines (Siniatchkin et al., 2000) in susceptible individuals.
SCP positivity is associated with increased cortical inhibition. Higher-than-expected positive SCPs have been noted in children with elevated blood lead levels (PbB; Otto & Reiter, 1984). Children diagnosed with ADHD show deficient SCP self-regulation skills compared with controls (Heinrich et al., 2004). SCPs have been used to monitor the depth of anesthesia during surgical procedures (Sebel et al., 1997) because they appear to be excellent indicators of the arousal level.
Recent studies have used SCPs to evaluate various task-oriented responses. Birbaumer and colleagues have trained SCPs to reduce seizures (Daum et al., 1993; Kotchoubey et al., 1997, 1999, 2001, 2002), and other groups have applied SCP feedback training to improve ADHD (Heinrich et al., 2004; Strehl, 2004, personal communication) and schizophrenia (Schneider et al., 1992).
SCP feedback training appears to be an approach that targets general characteristics of arousal using a single measure, compared to other types of EEG training that often reward increases and/or decreases in certain combinations of frequencies to accomplish changes in arousal. SCP feedback may provide a less complex approach to training neuronal activity in the clinical setting, providing greater accessibility via clinician-supervised home training devices.
Most research to date has been conducted using the Cz electrode site. However, at least one investigation involved training left hemisphere language sites. This approach demonstrated improved word processing results following the negativity training condition and diminished performance following the positivity condition (Pulvermuller, 2000). Studying the effects of SCP training at other electrode sites would be interesting.
Some efforts have been to identify SCP values using multiple electrodes in a quantitative EEG assessment paradigm. Basile and colleagues (2004) used four 32-channel DC-coupled amplifiers to identify differences in SCP responses in schizophrenic patients compared with normal controls. They found significant variations in response patterns, with normal controls showing simply-organized positivity and negativity patterns, while schizophrenic patients showed much more complex, fragmented patterns of activation and inhibition.
There are only a few clinically available DC-coupled amplifiers capable of accurately monitoring SCP activity. An Internet search yielded several devices aimed at the research institution market with correspondingly high prices and a couple of other devices with prices within a clinical practice’s reach. A new 32-channel DC-coupled data acquisition device for quantitative EEG assessments has also recently been released.
One potential attraction of using a DC amplifier is the capability of monitoring and/or training both SCPs and typical EEG frequencies. This is because DC amplifiers are optimized for SCP and have the capacity to record faster frequencies. This is particularly true for amplifiers with better analog-to-digital (AD) conversion characteristics (bit size, not sampling rate) because this allows them to record AC potentials without exceeding amplifier capabilities, which can be a problem in an amplifier without capacitors at the input stages.
Higher analog-to-digital (A/D) conversion values (more bits of data per sample) allow newer DC amplifiers to process EEG at a much lower voltage while retaining a high degree of resolution for signals that are often in the millivolt range (compared to microvolt values for most EEG signals).
The training of SCP shifts is a fairly new endeavor. Much remains to be learned about the effects of training both the positivity and negativity conditions at various electrode sites for individuals with various presenting concerns and specific neurophysiological characteristics.
This author's recent, brief clinical experiences suggest that training SCP using new, more accurate amplifiers may result in more pronounced changes occurring more quickly. This occurred on several occasions, even when using previously well-tested protocols alternating 8- to 10-second trials of both the positivity and negativity conditions. Thus, it will be important to develop protocols with more specificity and flexibility to meet the needs of non-homogeneous client populations that also consider changes in equipment and software characteristics that may affect the rate of skill acquisition and subsequent outcomes.
The author would like to thank Ute Strehl of the University of Tubingen in Germany, David Sever of Mind Alive, LTD in Canada, and Erwin Hartsuiker of Mind Media BV in the Netherlands for technical assistance in preparing this article.
NEUROPLASTICITY (LTD AND LTP)
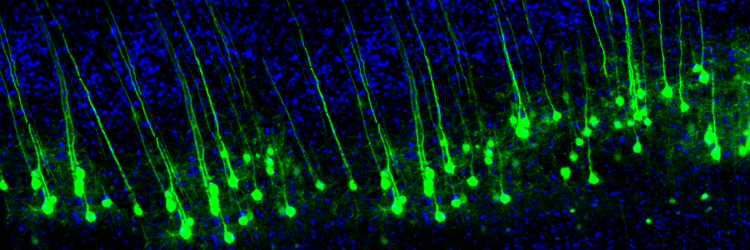
Neuroplasticity, the remodeling of neurons and neural networks with experience, is responsible for learning and memory. Memory storage involves the remodeling of neurons in terms of synaptic transmission, interneuron modulation, formation of new synapses, and rewiring of neural pathways (Bear, Connors, & Paradiso, 2020). Animal studies have shown that operant conditioning can induce astrogliogenesis (creation of new astrocytes) and neurogenesis (creation of new neurons) in structures like the medial prefrontal cortex and hippocampus (Rapanelli, Frick, & Zanutto, 2011).
The graphic is from synapse remodeling research by the Zuo Laboratory, MCD Biology, UCSC.
Neuroplasticity appears to involve a simple rule: when some synapses strengthen, adjacent synapses weaken to prevent overload due to increased input. A protein called Arc is crucial to this process (El-Boustani et al., 2018).
Neurofeedback, which involves the operant conditioning of CNS electrical activity, would be impossible without neuroplasticity. In neurofeedback, clients may learn to change the activity of local, regional, and global cortical resonant loops and the connectivity between brain regions (Collura, 2014; Thompson & Thompson, 2016).
To learn more about neuroplasticity, view the Khan Academy video Neuroplasticity.
Long-Term Depression and Long-term Potentiation
Two of the diverse processes involved in neuroplasticity are long-term depression and long-term potentiation.
In long-term depression (LTD),
synaptic transmission that coincides with slight depolarization of the postsynaptic neuron weakens a synapse due to pre- and postsynaptic changes. Relatively low-frequency stimulation of afferent neurons reduces the magnitude of their response to future stimulation
In long-term potentiation (LTP),
synaptic transmission that coincides with strong depolarization of the postsynaptic neuron strengthens a synapse due to pre- and postsynaptic changes. Strong stimulation of afferent neurons results in a stable and persistent (weeks or more) increase in synaptic effectiveness. LTP involves diverse changes, including creating new synapses, enhancing previous synapses, and building new dendritic branches and spines (Breedlove & Watson, 2020).
40-Hz rhythm: gamma rhythm hypothesized to be associated with feature binding
(linking an apple's color to its shape) and attributed to the neocortex and thalamocortical neurons.
acetylcholine: an amine neurotransmitter that binds to nicotinic and muscarinic
ACh receptors.
acetylcholine esterase (AChE): the enzyme that deactivates ACh.
AChE-R: an abnormal form of acetylcholine esterase (AChE) may render
dendrites with acetylcholine receptors more excitable when stressed.
action potential: a propagated electrical signal that usually starts at a neuron’s
axon hillock and travels to presynaptic axon terminals.
adenylate cyclase: at a metabotropic receptor, an enzyme that transforms ATP
into the second messenger cyclic AMP.
afferent: a neuron that transmits sensory information towards the central
nervous system or from one region to another.
allocortex: cortex that contains three or four layers and is comprised of the olfactory system and hippocampus.
all-or-none law: once an action potential is triggered in an axon, it is
propagated, without decrement, to the end of the axon. The amplitude of the action potential is unrelated to the
intensity of the stimulus that triggers it.
alpha blocking: arousal and specific forms of cognitive activity may reduce
alpha amplitude or eliminate it entirely while increasing EEG power in the beta range.
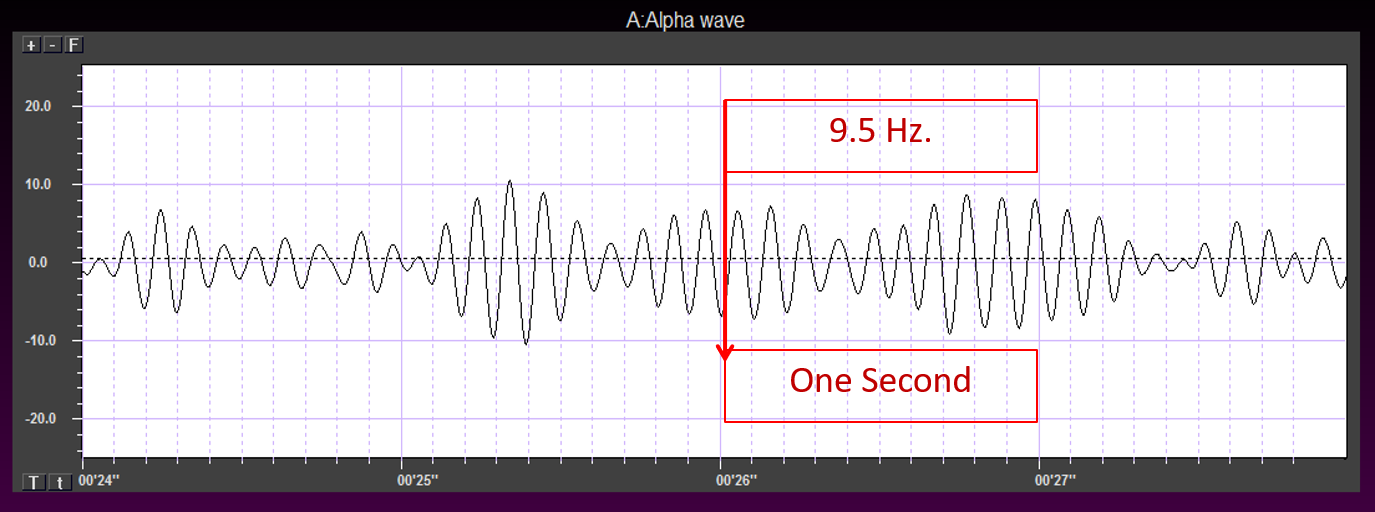
alpha rhythm: 8-12-Hz activity that depends on the interaction between rhythmic burst firing by a subset of thalamocortical (TC) neurons that are linked by gap junctions and rhythmic inhibition by widely distributed reticular nucleus neurons. Researchers have correlated the alpha
rhythm with "relaxed wakefulness." Alpha is the dominant rhythm in adults and is located posteriorly. The alpha rhythm may be divided into alpha 1 (8-10 Hz) and alpha 2 (10-12 Hz).
alpha spindles: regular bursts of alpha activity.
alpha-subunit: a subunit of a G protein associated with the neuron membrane that breaks away to activate enzymes within the neuron when a ligand binds to a metabotropic receptor.
amino acid neurotransmitters: the oldest family of transmitters. These molecules
bind to ionotropic and metabotropic receptors, so they transmit information and modulate neuronal activity. In the
brain, most synaptic communication is accomplished by glutamate (generally excitatory) and GABA (generally
inhibitory).
AMPA (glutamate) receptors: ionotropic receptors which open sodium channels,
depolarize the neuron's membrane (producing an EPSP), and dislodge a Mg+ ion that blocks an adjacent NMDA
(glutamate) receptor's calcium channel. AMPA receptors are responsible for most activity at glutamatergic
synapses.
amplitude: the energy or power contained within the EEG signal measured in
microvolts or picowatts.
amygdala: the limbic system structure that participates in evaluating whether
stimuli are salient (rewarding or threatening), establishing unconscious emotional memories, learning conditioned emotional responses,
and producing anxiety and fear responses.
anion: a negative ion, for example, chloride (Cl-).
anterior: near or toward the front of the head, for example, the anterior
cingulate.
anterior cingulate: a division of the prefrontal cortex that plays a vital
role in attention and is activated during working memory. It mediates emotional and physical pain, and has
cognitive (dorsal anterior cingulate) and affective (ventral anterior cingulate) conflict-monitoring
components.
anterior commissure: a bundle of nerve fibers that crosses the midline and
connects the left and right temporal lobes and the hippocampus and amygdala.
apical dendrite: a dendrite that arises from the top of the pyramid and extends
vertically to layer 1 of the neocortex.
arousal: a process that combines alertness and wakefulness, produced by at least
five neurotransmitters, including acetylcholine, histamine, hypocretin, norepinephrine, and serotonin.
aspinous (smooth) neurons: neurons without dendritic spines that are believed to be inhibitory.
astrocytes: star-shaped glial cells that communicate with and support neurons
and help determine whether synapses will form.
asynchronous waves: neurons depolarize and hyperpolarize independently.
ATP: energy source for a neuron’s sodium-potassium transporters.
autoreceptors: metabotropic receptors that can be located on the membrane of
any part of a neuron. They detect neurotransmitters the neuron releases, generate IPSPs that inhibit the
neuron from reaching the excitation threshold, and regulate internal processes like transmitter synthesis and release through the
second messenger system.
axoaxonic synapses: junctions between two axons that do not affect the
generation of an action potential, only the amount of neurotransmitter distributed.
axodendritic synapses: junctions between axons and dendrites that determine
whether the axon hillock will initiate an action potential.
axon: long, cylindrical structures that convey information from the soma to
the terminal buttons. An axon also transports molecules in both directions along the outer surface of protein
bundles called microtubules.
axon hillock: a swelling in the cell body where a neuron integrates the
messages it has received from other neurons and decides whether to fire an action potential.
axon terminal: buds located on the ends of axon branches that form synapses and release neurochemicals to other neurons.
axonal varicosity: a swelling in an axon wall that releases
neurotransmitters through the wall via volume transmission.
axoplasmic transport: the movement of molecules in both directions along the outer
surface of protein bundles called microtubules.
basal dendrite: a dendrite that horizontally branches out from the 30 μm
base of the pyramid through the layer where the neuron resides.
basal ganglia: forebrain structures consisting of an egg-shaped nucleus
that contains the putamen and globus pallidus and a tail-shaped structure called the caudate,
which together are responsible for the production of movement. The basal ganglia have also been implicated in
obsessive-compulsive disorder, Parkinson’s disease, and Huntington’s chorea.
beta rhythm: 12-38-Hz activity associated with arousal and attention
generated by brainstem mesencephalic reticular stimulation that depolarizes neurons in both the thalamus and
cortex. The beta rhythm can be divided into multiple ranges: beta 1 (12-15 Hz), beta 2 (15-18 Hz), beta 3 (18-25 Hz), and beta 4 (25-38 Hz).
bilateral synchronous slow waves: a pathological sign observed in drowsy
children. When detected in alert adults, intermittent bursts of high amplitude slow waves may signify gray
matter lesions in deep midline structures.
cation: positive ion, for example, sodium (Na+).
caudal: away from the front of the head.
cell body or soma: part of a neuron that contains machinery for cell life processes and receives and integrates EPSPs and IPSPs from axons generated by axosomatic synapses (junctions between axons and somas). The cell body of a typical neuron is 20 μm in diameter, and its spherical nucleus, which contains
chromosomes comprised of DNA, is 5-10 μm across.
central nucleus of the amygdala: nucleus that orchestrates the nervous
system's response to important stimuli by activating circuits in the brainstem (autonomic arousal) and the basal
ganglia and periaqueductal gray (defensive behavior).
cerebral cortex: the layer of gray matter that covers the cerebral
hemispheres. The cerebral cortex consists of gray matter and white matter.
chemical synapses: junctions between neurons that transmit molecules across
gaps of less than 300 angstroms. Neurons use chemical synapses to produce short-duration (millisecond) and long-duration
(seconds to hours) changes in the nervous system. Chemical synapses are capable of more extensive communication and initiating more diverse and long-lasting changes than electrical synapses.
classical routes for EEG activation: specific sensory pathways like the visual
(retina to the visual cortex), auditory (cochlea to the auditory cortex), and somatosensory (chemoreceptors and
mechanoreceptors to the somatosensory cortex) systems. Increased transmission of information through these
pathways desynchronizes EEG activity in the cortical regions to which these afferent neurons project, as
specialized circuits of neurons independently process this information.
commissures: axon tracts. The left and right hemispheres communicate using the
corpus callosum, anterior commissure, and posterior commissure.
complex: a sequence of waves.
COMT: a degrading enzyme that only targets the catecholamines dopamine and
norepinephrine.
contingent negative variation (CNV): a steady, negative shift in potential (15
microvolts in young adults) detected at the vertex. This slow cortical potential may reflect expectancy,
motivation, intention to act, or attention. The CNV appears 200-400 ms after a warning signal (S1), peaks within
400-900 ms, and sharply declines after a second stimulus that requires the performance of a response (S2).
continuous irregular delta: slow waves produced by white matter lesions seen
in disorders like multiple sclerosis.
contralateral: structures that are located on opposite sides of the body. For
example, neurons in the left primary motor cortex control muscles on the right side of the body.
corpus callosum: the largest commissure that connects the left and right
frontal, parietal, and occipital lobes.
corticothalamic network: a unified network that generates diverse types of brain
rhythms grouped by slow cortical oscillations.
cyclic AMP: a second messenger that moves about the neuron, activating other
enzymes. Protein kinase A, which controls the excitability of ion channels, is a crucial enzyme target of
cyclic AMP. Cyclic AMP also travels to the nucleus, where it can regulate gene expression.
Dale's principle: incorrect view that a neuron can only release one
neurotransmitter. They often release two to four.
delta rhythm: 0.05-3 Hz oscillationsgenerated
by thalamocortical neurons during stage 3 sleep.
dendrite: a branched structure designed to receive messages from other neurons via axodendritic synapses (junctions between axons and dendrites), determining whether the axon hillock will initiate an action potential.
dendritic spines: protrusions on the dendrite shaft where axons typically form
axodendritic synapses.
dendrodendritic synapses: junctions between dendrites that communicate
chemically across synapses and electrically across gap junctions.
depolarization: to make the membrane potential less negative by making the inside
of the neuron less negative with respect to its outside.
diffusion: the distribution of molecules from areas of high concentration to
low concentration.
diphasic wave: a wave that contains both a negative and positive deflection from
the baseline.
dipole: the electrical field generated between the sink (where current enters
the neuron ) and the source (place at the other end of the neuron where current leaves), which may be located
anywhere along the dendrite.
dominant frequency: the EEG frequency with the most significant amplitude.
dopamine: a monoamine neurotransmitter exerts its postsynaptic effects on at
least six receptors linked to G proteins. This means that dopamine functions as a neuromodulator.
The two families include D1 (D1 and D5) and D2 (D2A, D2B, D3, and D4).
dorsal: toward the upper back or head.
dorsolateral prefrontal cortex: the left dorsolateral prefrontal cortex is
concerned with approach behavior and positive affect. It helps us select positive goals and organizes and
implements behavior to achieve these goals. The right dorsolateral prefrontal cortex organizes withdrawal-related
behavior and negative affect and mediates threat-related vigilance. It plays a role in working memory for object
location.
D-serine: a neurotransmitter that binds to the glycine site on the NMDA receptor
to trigger calcium entry into a dendritic spine when glutamate binds to its site, resulting in a large, prolonged
increase in intracellular calcium.
dual-action antidepressants: medications that activate 5-HT1 receptors to
produce antidepressant and anxiolytic effects, while they blockade 5-HT2 (agitation, restlessness, and sexual
dysfunction) and 5-HT3 (nausea, headache, and vomiting) receptors to minimize their side effects.
EEG activity: single wave or successive waves.
EEG power: the signal energy in the EEG spectrum. Most EEG power falls
within the 0-20 Hz frequency range. EEG power is measured in microvolts or picowatts.
efferent: motoneuron that transmits information towards the periphery.
electrical synapse: symmetrical synapse where neurons communicate information
bidirectionally across gap junctions between adjacent membranes using ions. Transmission across electrical
synapses is instantaneous, compared with the 10-ms or longer delay in chemical synapses. The rapid information
transmission that characterizes electrical synapses enables large circuits of distant neurons to synchronize their
activity and simultaneously fire.
electroencephalogram (EEG): the voltage difference between at least two
electrodes, where at least one electrode is located on the scalp or inside the brain. The EEG is a recording of
EPSPs and IPSPs that occur primarily in dendrites in pyramidal cells located in macrocolumns, several millimeters in
diameter, in the upper cortical layers.
electrostatic pressure: the attractive or repulsive force between ions that
moves them from one region to another.
entorhinal cortex: a structure located in the caudal region of the temporal lobe and receives pre-processed sensory information from all modalities and reports on cognitive operations. The
entorhinal cortex provides the main input to the hippocampus, and is involved in memory consolidation, spatial
localization, and provides input into the septohippocampal system that may generate the 4-7 Hz theta rhythm.
enzymatic deactivation: the process in which an enzyme breaks a neurotransmitter
apart into inactive fragments. For example, acetylcholine transmission is ended by the enzyme acetylcholine
esterase (AChE). Deactivating enzymes located in the synaptic cleft degrade a neurotransmitter molecule when it detaches from its binding site.
evoked potential: an event-related potential (ERP) elicited by external sensory
stimuli (auditory, olfactory, somatosensory, and visual). An evoked potential has a negative peak at 80-90 ms and
a positive peak around 170 ms following stimulus onset. The orienting response ("What is it?") is a sensory
ERP. The N1-P2 complex in the auditory cortex of the temporal cortex reveals whether an uncommunicative person can
hear a stimulus.
excitatory postsynaptic potential (EPSP): a brief positive shift in a
postsynaptic neuron's potential produced when neurotransmitters bind to receptors and cause positive
sodium ions to enter the cell. An EPSP pushes the neuron towards the threshold of excitation when it can initiate
an action potential.
exocytosis: the process of neurotransmitter release. When an action potential
arrives and depolarizes the terminal button, calcium ions enter the terminal button from the extracellular fluid.
Calcium binds with clusters of protein molecules that connect the vesicles to the presynaptic membrane. The
clusters move apart, forming a hole through both membranes called a fusion pore, and the neurotransmitter leaves the
terminal button for the synaptic cleft or extracellular fluid.
exogenous ERP: an event-related potential (ERP) elicited by external sensory
stimuli (auditory, olfactory, somatosensory, and visual).
explicit learning: behavioral changes that occur with our conscious awareness
that require processing by the hippocampus.
extracellular dipole layers: macrocolumns of pyramidal cells, which lie
parallel to the surface of the cortex, send opposite charges towards the surface and the deepest of the 5-7 layers
of cortical neurons.
extracellular fluid: the fluid surrounding a neuron.
fast cortical potentials: EEG rhythms that range from 0.5 Hz-100 Hz. The main
frequency ranges include delta, theta, alpha, sensorimotor rhythm, and beta.
feature binding: the process of linking information to perceptual objects
(linking an apple's color to its shape) that may involve the 40-Hz rhythm.
fissures: deep grooves, for example, the lateral fissure.
focal waves: EEG waves that are detected within a limited area of the scalp,
cerebral cortex, or brain.
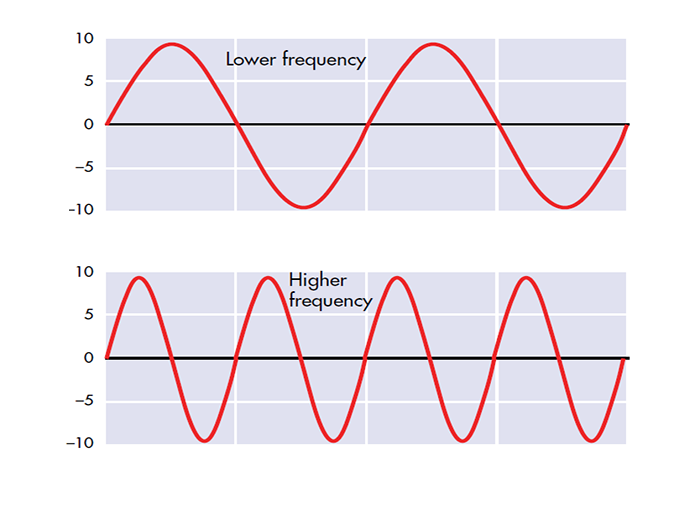
frequency: the number of cycles completed each second expressed in hertz (Hz).
frequency synchrony: when identical EEG frequencies are detected at two or
more electrode sites. For example, 12 Hz may be simultaneously detected at O1-A1 and O2-A2.
frontal lobes: the most anterior cortical lobes of the brain that are divided
into the motor cortex, premotor cortex, and prefrontal cortex.
fusion pore: a hole through a vesicle and presynaptic membrane that allows
neurotransmitter to leave the terminal button for the synaptic cleft or extracellular fluid.
G protein: a protein located inside a neuron’s membrane next to a
metabotropic receptor that is activated when the receptor binds a ligand. An alpha-subunit of the G protein then
breaks away to perform actions within the cell.
GABA: an amino acid that is often inhibitory and that may be the most
important inhibitory neurotransmitter in the brain. There are several types of GABA receptors, each of which
produces inhibition differently.
gamma rhythm: EEG activity frequencies above 30 or 35 Hz. Frequencies from 25-70 Hz are called low gamma, while those above 70 Hz represent high gamma.
gap junction: narrow space between two cells bridged by connexons (protein channels) that allow ions to travel between them rapidly.
generalized asynchronous slow waves: waves that are seen in sleepy children
and those with elevated temperatures. These waves may indicate degenerative disease, dementia, encephalopathy, head injury, high fever, migraine, and Parkinson's disease in adults.
glial cells: nonneural cells that guide, insulate, and repair neurons and
provide structural, nutritional, and information-processing support. Glial cells generate slow cortical
potentials (SCPs). Glial cells include astrocytes, microglia, oligodendrocytes, radial glial cells, and Schwann
cells.
glutamate: an amino acid that is often excitatory and that may be the primary
excitatory neurotransmitter in the brain. Its receptors are found on the surface of almost all neurons. There are
at least 13 different receptors for glutamate, 5 ionotropic and 8 metabotropic. Most presynaptic neurons in the
brain excite postsynaptic neurons via ionotropic glutamate receptors in the postsynaptic membrane. Metabotropic
glutamate receptors may play a regulatory function, either augmenting or suppressing the activation of ionotropic
glutamate receptors.
glycine: an amino acid that is often inhibitory and has a binding
site on the NMDA receptor.
gray matter: brain tissue that looks grayish brown and comprises cell
bodies, dendrites, unmyelinated axons, glial cells, and capillaries.
gyrus: ridge of cortex demarcated by sulci or fissures, for example, the
precentral gyrus.
hertz (Hz): the unit of frequency, an abbreviation for cycles per second.
hippocampus: a limbic structure located in the medial temporal lobe
involved in 4-7 Hz theta activity, control of the endocrine system’s response to stressors, formation of
explicit memories, and navigation. Cortisol binding to this structure disrupts these functions, interferes with
creating new neurons, and harms and kills hippocampal neurons.
hubs: highly centralized nodes through which other node pairs communicate;
hubs allow efficient communication.
hyperpolarize: a negative shift in membrane potential (the inside becomes more
negative with respect to the outside) due to the loss of positive ions or gain of negative ions.
inhibitory postsynaptic potential (IPSP): a brief negative shift in a postsynaptic neuron's potential produced when cations like potassium leave a neuron or anions (negative ions) like chloride enter a neuron, which hyperpolarize the cell. An IPSP pushes the neuron away from its excitation
threshold.
integration: the addition of EPSPs and IPSPs at the axon hillock. Neurons sum
EPSPs and IPSPs over their surface in spatial integration and over milliseconds in temporal
integration to raise the membrane from its resting potential to the excitation threshold. EPSPs and IPSPs
last from 15-200 ms, while action potentials occur in 1-2 ms.
interneurons: neurons that receive input from and distribute output to other
neurons. They have short processes and are confined to the central nervous system. They provide the integration
required for decisions, learning and memory, perception, planning, and movement.
intracellular fluid: the watery cytoplasm contained within a neuron.
ion: a charged atom or molecule with a positive or negative charge. Positive
ions are called cations, and negative ions are called anions.
ionotropic receptor: receptor protein that contains a binding site for a
ligand and an ion channel that opens when the neurotransmitter attaches to this site.
ipsilateral: structures that are located on the same side of the body. For
example, the left olfactory bulb distributes axons to the left hemisphere.
irregular waves: successive waves that constantly alter their shape and
duration.
kappa rhythm: bursts of alpha or theta and is detected over the temporal lobes
of subjects during cognitive activity.
lambda waves: saw-toothed transient waves from 20-50 μV in amplitude and
100-250 ms in duration detected over the occipital cortex during wakefulness. These positive deflections are
time-locked to saccadic movements and observed during visual scanning, as during reading.
lateral: to the side, away from the center, as in the lateral geniculate
nucleus.
lateral nucleus of the amygdala: a nucleus that processes sensory information
and distributes it throughout the amygdala.
lateralized waves: waves that are primarily detected on one side of the scalp
and that may indicate pathology.
Layers I-III: cortical layers that receive corticocortical afferent fibers
that connect the left and right hemispheres.
Layer III: the cortical layer that is the primary source of corticocortical efferent
fibers.
Layer IV: the cortical layer that is the primary destination of
thalamocortical afferents and intra-hemispheric corticocortical afferents.
Layer V: the cortical layer that is the primary origin of efferent fibers that
target subcortical structures that have motor functions.
Layer VI: the cortical layer that projects corticothalamic efferent fibers to
the thalamus, which, together with the thalamocortical afferents, creates a dynamic and reciprocal relationship
between these two structures.
left dorsolateral prefrontal cortex: the division of the prefrontal cortex
concerned with approach behavior and positive affect. It helps us select positive goals and organizes and
implements behavior to achieve these goals.
local field potential: the aggregate effect of the firing of the interconnected pyramidal neurons within the cortical columns plus additional mechanisms like glial cell modulation of the cortical electrical gradient.
local synchrony: synchrony that occurs when high-amplitude EEG signals are
produced by the coordinated firing of cortical neurons.
localized slow waves: waves that may indicate a transient ischemic attack
(TIA) or stroke, migraine, mild head injury, or tumors above the tentorium. Deep lesions result in bilateral or
unilateral delta.
locus coeruleus: the noradrenergic branch of the ascending reticular activating
system, which is responsible for vigilance. Subnormal norepinephrine transmission may contribute to ADHD.
long-latency potentials: potentials that have extended latencies following
stimulus onset, for example, P300 and N400 ERPs.
long-term depression (LTD): a persistent decrease in synaptic strength
following low-frequency stimulation.
long-term potentiation (LTP): a persistent increase in synaptic strength
following high-frequency stimulation.
macrocolumns: circuits of cortical pyramidal neurons several millimeters in diameter
that create extracellular dipole layers parallel to the surface of the cortex that send opposite charges towards
the surface and the deepest of the 5-7 layers of cortical neurons. Since the pyramidal neurons are all
aligned with the cortical surface, the postsynaptic potentials at cells within the same macrocolumn add together. This summation occurs
because they share the same charge and the macrocolumns fire synchronously.
medial: toward the center of the body, away from the side. For example, the
medial geniculate nucleus.
medial prefrontal cortex: the division of the prefrontal cortex that integrates
cognitive-affective information and helps control the hypothalamic–pituitary–adrenal (HPA) axis during
emotional stress.
membrane potential: a neuron’s electrical charge created by a difference
in ion distribution within and outside the neuron. A typical resting potential is about -70 mV
(thousandths of a volt), since the inside of a resting axon is more negatively charged than the outside.
mesocortical neurons: dopaminergic neurons that project from the ventral
tegmental area of the midbrain to the prefrontal cortex and excite prefrontal cortical neurons that control
working memory, planning, and strategy preparation for problem-solving. Underactivity in this pathway is
associated with the negative symptoms of schizophrenia-like attentional deficits.
metabotropic receptors: include all G protein-linked receptors located on
neurons, including autoreceptors. Neurotransmitters that bind to G protein-linked receptors are often called
neuromodulators. Metabotropic receptors, which indirectly control the cell's operations, expend energy, and
produce slower, longer lasting, and more diverse changes than ionotropic receptors. Their effects can last several
seconds, instead of milliseconds, because of the long-lived activity of G proteins and cyclic AMP.
microglia: microscopic glial cells that participate in the immune response.
microtubules: hollow cylindrical protein bundles that are involved in
axoplasmic transport.
modulating effects: neuromodulators like the monoamines alter the performance
of diffuse networks of target neurons by indirectly controlling cellular operations when they bind to metabotropic
receptors.
module: a set of interconnected nodes in a neural network.
monoamine neurotransmitters: amine neurotransmitters that include
dopamine, norepinephrine, epinephrine (catecholamines), and serotonin (indoleamine). These neurotransmitters
are released using volume transmission and generally have modulating effects, altering the performance of diffuse
networks of target neurons.
monoamine oxidase (MAO): an enzyme that degrades and inactivates the monoamine
neurotransmitters dopamine, norepinephrine, and serotonin.
monoamine oxidase inhibitors (MAOIs): antidepressant drugs that interfere with
MAO's breakdown of monoamines and increase monoamine availability to treat clinical depression.
monophasic wave: either a single negative (upward) or positive (downward)
deflection from baseline.
motor cortex: the subdivision of the frontal lobe located in the
precentral gyrus and guides fine motor coordination (like writing).
motor ERPs: event-related potentials detected over the primary motor cortex
(precentral gyrus) during movement. Their amplitude is proportional to the force and rate of skeletal muscle
contraction.
motor neurons: efferent neurons that convey commands to glands, muscles, and
other neurons.
movement-related potentials (MRPs): slow cortical potentials that occur at 1
second as subjects prepare for unilateral voluntary movements. MRPs are distributed bilaterally with maximum
amplitude at Cz. The supplementary motor area and primary motor and somatosensory cortices primarily generate
these potentials.
mu rhythm: 7-11-Hz waves resemble wickets and appear as several-second trains over central or centroparietal sites (C3 and C4).
multiple spike-and-slow-wave complex: multiple spikes associated with at least
one slow wave.
muscarinic receptors: metabotropic ACh receptors that are stimulated by
muscarine and blocked by atropine. Muscarinic receptors control smooth muscle and predominate in the CNS. In the
CNS, muscarinic receptors help mediate learning, memory, attention, arousal, EEG, and postural control.
myelinated axons: axons that are insulated by myelin by oligodendrocytes in
the central nervous system and Schwann cells in the peripheral nervous system.
N1-P2: a sensory event-related potential in the auditory cortex of the
temporal cortex that reveals whether an uncommunicative person can hear a stimulus.
N400 potential: an event-related potential (ERP) elicited when we encounter
semantic violations like ending a sentence with a semantically incongruent word ("The handsome prince married
the beautiful fish"), or when the second word of a pair is unrelated to the first (BATTLE/GIRL).
negative SCPs: slow cortical potentials produced by glial cells that increase
the probability of neuron firing.
neuroaxis: an imaginary line that runs centrally through the central nervous
system (CNS) from the front of the prefrontal cortex to the base of the spinal cord.
neuromodulator: a neurochemical that modifies the effect of neurotransmitters
through mechanisms like binding to metabotropic receptors.
neuron: a nerve cell that is the fundamental anatomical unit of the nervous
system.
nicotinic ACh receptor: an ionotropic receptor that is stimulated by nicotine and
blocked by curare. They are mainly found in the PNS on skeletal muscles. At CNS axoaxonic synapses, they produce
presynaptic facilitation (increase neurotransmitter release). In the CNS, nicotinic receptors help regulate
cortical blood flow, anxiety reduction, and decision-making.
nigrostriatal pathway: dopaminergic pathway from the substantia nigra to the
basal ganglia (caudate nucleus and putamen) that controls movement. The nigrostriatal pathway is progressively
destroyed in Parkinson’s disease.
nitric oxide: a gaseous retrograde transmitter that is involved in long-term
potentiation (LTP).
NMDA (glutamate) receptors: ligand-gated and voltage-gated glutamate receptors
that bind the glutamate agonist NMDA. NMDA receptors play an essential role in long-term potentiation (LTP).
node: vertex within a neural network.
nodes of Ranvier: gaps between myelinated axon segments where the axon
membrane is exposed to extracellular fluid and action potentials are regenerated by sodium ion entry.
norepinephrine: a monoamine neurotransmitter that exerts postsynaptic effects at
alpha and beta receptors, each with two subtypes. All norepinephrine receptors are G protein-linked. The
cell bodies of the core noradrenergic system are located in the locus coeruleus, a nucleus found in the
dorsal pons.
nucleus accumbens: a limbic structure that is a target of dopamine released by
the mesolimbic pathway. The nucleus accumbens plays a critical role in the reinforcement of diverse activities,
including ingestion of drugs like central nervous system stimulants.
occipital lobes: cortical lobes that are posterior to the parietal lobes. They
process visual information from the eyes in collaboration with the frontal, parietal, and temporal
lobes.
odd-ball stimulus: a meaningful stimulus that is different from others in a
series used to elicit the P300 potential. For example, a colored playing card in a series of monochrome
cards.
oligodendrocytes: glial cells that insulate adjacent axons within the brain and spinal cord of the central nervous system.
orbitofrontal cortex: the frontal lobe subdivision that is concerned with
affective evaluation. It decodes the punishment and reward value of stimuli and helps inhibit inappropriate
behavior. Phineas Gage's profound personality changes were produced by damage to this region.
orienting response: Pavlov’s "What is it?" reaction to stimuli
like the sound of a vase crashing that includes (1) increased sensory sensitivity, (2) head (and ear) turning
toward the stimulus, (3) increased muscle tone (reduced movement), (4) EEG desynchrony, (5) peripheral
constriction and cephalic vasodilation, (6) a rise in skin conductance, (7) heart rate slowing, and (8) slower,
deeper breathing.
P300 potential: an event-related potential (ERP) with a 300-900-ms latency. The largest amplitude positive peaks are located over the parietal lobe. The P300 potential may reflect an event’s
subjective probability, meaning, and transmission of information.
paralimbic cortex: a transitional region between neocortex and allocortex.
parietal lobes: cortical lobes posterior to the frontal lobes that are divided
into the primary somatosensory cortex (postcentral gyrus) and secondary somatosensory cortex. Their primary function
is to process somatosensory information like pain and touch. The right posterior parietal lobe helps guide
movements, locate objects in three-dimensional space, and create body boundaries.
phase: the degree to which the peaks and valleys of EEG waveforms
coincide.
phase synchrony: synchrony when identical EEG frequencies are detected at two
or more electrode sites, and the peaks and valleys of the EEG waveforms coincide. This is also called global
synchrony. For example, EEG training may produce phase-synchronous 12-Hz alpha waves at O1-A1 and O2-A2.
polarization: to make the membrane potential more negative by making the inside
of the neuron more negative with respect to its outside.
polyphasic (multiphasic) wave: a wave that contains two or more deflections of
opposite polarity from baseline.
positive SCPs: slow cortical potentials that are produced by glial cells that
decrease the probability of neuron firing.
posterior: near or toward the back of the head.
posterior commissure: axon tracts located below the corpus callosum that
connect the right and left diencephalon and mesencephalon.
precision: the number of voltage gradations or steps.
prefrontal cortex: the most anterior frontal lobe division and is subdivided
into dorsolateral, medial, orbitofrontal, and anterior cingulate regions responsible for executive
functions like attention and planning.
premotor cortex: the frontal lobe subdivision that is anterior to the motor cortex
and helps program head, trunk, and limb movements.
presynaptic facilitation: a modulatory process in which a neuron increases the
presynaptic neuron's neurotransmitter release by delivering a neurotransmitter that increases calcium ion entry
into its terminal button.
presynaptic inhibition: a modulatory process in which a neuron decreases
neurotransmitter release by reducing calcium ion entry.
primary somatosensory cortex (S1): the parietal lobe subdivision located at
the postcentral gyrus that processes information about touch and pain.
protein kinase A: an intracellular enzyme that controls the excitability of
ion channels and is a critical enzyme target of cyclic AMP.
raphe nuclei: serotonergic cell bodies in the midbrain, pons, and medulla
give rise to most of the brain’s serotonergic neurons.
rate law: the principle that neurons represent the intensity of a stimulus by
variation in the rate of axon firing.
readiness potential: slow-rising, negative potential (10-15 µV)
detected at the vertex before voluntary and spontaneous movement. This slow cortical potential precedes voluntary
movement by 0.5 to 1 second and peaks when the subject responds.
regular or monomorphic waves: successive waves with identical shapes.
Regular waves may resemble sine waves (sinusoidal) or maybe arched (resembling wickets) or saw-toothed
(asymmetrical and triangular).
resting potential: the membrane potential of a neuron when it is not influenced by
messages from other neurons.
reticular formation: a network of 90 nuclei within the central brainstem from the lower medulla to the upper midbrain. The reticular formation sends axons to the spinal cord, thalamus, and cortex, contributing to diverse functions like neurological reflexes, muscle tone and movement, attention, arousal, and sleep.
reuptake: the primary method that neurons terminate the action of
neurotransmitters. Reuptake transporters located in terminal buttons and astrocytes remove neurotransmitters from
the synaptic cleft.
reward deficiency syndrome: Blum’s hypothesis that an abnormal form of
the A1 allele is present in most severe alcoholics and results in defective D2 receptors. Reduced D2 receptor
activity may reduce the activation of the nucleus accumbens and hypothalamus and result in dysphoria, drug
craving, and compulsive drug-seeking and abuse.
right dorsolateral prefrontal cortex: the division of the prefrontal cortex that
organizes withdrawal-related behavior and negative affect and mediates threat-related vigilance. It plays a role
in working memory for object location.
rostral: toward the front of the head.
saltatory conduction: action potential conduction in myelinated axons in which
action potentials jump from node to node for 200 times greater speed.
sampling rate: the number of measurements per second (Hz).
Schwann cells: glial cells that provide myelin for single PNS axons and facilitate axonal regeneration following damage.
secondary somatosensory cortex (S2): a region of the parietal lobe that receives
somatosensory information from the primary somatosensory cortex (S1).
sensorimotor rhythm (SMR): EEG rhythm that ranges from 12-15 Hz and is located
over the sensorimotor cortex (central sulcus). The waves are synchronous. The sensorimotor rhythm is associated
with the inhibition of movement and reduced muscle tone. The SMR is generated by "ventrobasal relay cells in
the thalamus and thalamocortical feedback loops."
sensorimotor system: in Sterman’s model, ascending pathways that convey
information about touch and proprioception to the thalamus, the thalamus and its thalamic projections to the
sensorimotor cortex, and the sensorimotor cortex, and its efferent fibers.
sensory event-related potentials (ERPs): event-related potentials evoked by external sensory
stimuli (auditory, olfactory, somatosensory, and visual). These evoked potentials or exogenous ERPs have a
negative peak at 80-90 ms and a positive peak around 170 ms following stimulus onset. These changes in brain activity in response to
specific stimuli. ERPs can be detected throughout the cortex. Investigators monitor
ERPs by placing electrodes at locations like the midline (Fz, Cz, and Pz). A computer analyzes a subject's EEG
responses to the same stimulus or task over many trials to subtract random EEG activity. ERPs always have the same
waveform morphology. Their negative and positive peaks occur at regular intervals following the stimulus.
sensory neurons: neurons specialized for sensory intake. They are called
afferent because they transmit sensory information towards the central nervous system (brain and spinal cord).
septal nuclei: in Sieb’s model, when the prefrontal cortex receives
information about high-priority environmental events, it signals cell bodies in the septum to induce a beta
rhythm in the hippocampus to remove its inhibition of vigilance centers.
septohippocampal system: a subcortical circuit from the septum to hippocampus
that contributes to 4-7 Hz theta activity.
septum: a limbic structure that contains several nuclei involved in emotion and
addiction and control of aggressive behavior.
sharp transients: a sequence that contains several sharp waves.
sharp waves: waves that resemble spikes with a pointed peak with a longer
70-200-ms duration.
sink: a site where current enters the neuron. Positive sodium ion entry into a
neuron creates an active sink, represented by -ve.
slow cortical potentials (SCPs): gradual changes in the membrane potentials of
cortical dendrites that last from 300 ms to several seconds. These potentials include the contingent negative
variation (CNV), readiness potential, movement-related potentials (MRPs), and P300 and N400 potentials. SCPs
modulate the firing rate of cortical pyramidal neurons by exciting or inhibiting their apical dendrites. They
group the classical EEG rhythms using these synchronizing mechanisms.
sodium (Na+) ions: positive ions that enter a neuron during EPSPs
and action potentials.
sodium-potassium transporters: pumps that are powered by ATP and that exchange
three sodium for two potassium ions.
soma or cell body: the region of a neuron that contains machinery for cell life processes and receives and integrates EPSPs and IPSPs from axons generated by axosomatic synapses (junctions between axons and somas). The cell body of a typical neuron is 20
μm in diameter, and its spherical nucleus, which contains chromosomes comprised of DNA, is 5-10 μm
across.
source: the place at the end of the neuron opposite of the sink where current
leaves, represented by +ve. The extracellular area surrounding the source becomes electrically
positive.
spatial summation: the addition of EPSPs and IPSPs over a neuron’s
surface.
spike: a negative transient with a pointed peak at conventional paper speeds,
20-70-ms duration, and 40-100 μV amplitude; rare dendritic action potentials.
spike-and-slow-wave complex: a spike followed by a higher amplitude
slow wave at 3 Hz. In an absence seizure, the amplitudes are very high (e.g., 160 μV).
spindle waves: waves that originate in the thalamus and occur during
unconsciousness and stage II sleep.
spiny neurons: neurons with dendritic spines that are usually excitatory.
striatal: basal ganglia (caudate nucleus and putamen).
Stroop test: cognitive monitoring task where color and names conflict.
substantia nigra: midbrain structure that projects to the basal ganglia
(caudate nucleus and putamen) to control movement and that is progressively destroyed in Parkinson’s
disease.
sulcus: a shallow groove in the surface of the cerebral hemisphere, for
example, the central sulcus.
synapse-associated polyribosome complexes (SPRCs): organelles with dendrites
that can produce proteins that allow rapid remodeling of synapses. A polyribosome complex consists of several
ribosomes bound to messenger RNA (mRNA). SPRCs represent one mechanism underlying synaptic plasticity.
synaptic cleft: 20-40-nm fluid-filled gap between presynaptic and postsynaptic
structures.
synchronous: adverb meaning that groups of neurons depolarize and
hyperpolarize simultaneously.
synchrony: the coordinated firing of pools of neurons. EEG signals can display
local synchrony, frequency synchrony, and phase synchrony.
telencephalon: the frontal subdivision of the forebrain, including the
cerebral cortex, basal ganglia, and limbic system.
temporal summation: the addition of EPSPs and IPSPs over time. Summation is
more effective when postsynaptic potentials are generated more closely in time.
temporal lobes: lobes separated from the rest of the cortical lobes by the
Sylvian fissure. The temporal lobes process hearing, smell, and taste information and help us understand spoken
language and recognize visual objects and faces. The amygdala and hippocampus, which lie beneath the temporal
cortex, play crucial roles in emotion, declarative, emotional, and working memory, and navigation.
terminal buttons: buds located on the ends of axon branches that form synapses
and release neurochemicals to other neurons. They contain vesicles that store neurotransmitters for release when
an action potential arrives. A terminal button’s presynaptic membrane may possess reuptake transporters that
return neurotransmitters from the synapse or extracellular space for repackaging.
thalamus: forebrain structure above the hypothalamus that receives, filters,
and distributes most sensory information. The thalamus contains neurons that can block or relay ascending sensory
information. When these thalamic neurons rhythmically fire, this blocks the transmission of information to the cortex.
When they depolarize in response to sensory information, this integrates and transmits this information to the
cortex. Inputs to the thalamus determine whether these neurons block or relay sensory information.
theta rhythm: 4-8-Hz rhythms generated a cholinergic septohippocampal system
that receives input from the ascending reticular formation and a noncholinergic system that originates in the
entorhinal cortex, which corresponds to Brodmann areas 28 and 34 at the caudal region of the temporal lobe.
threshold of excitation: the membrane potential at which an axon initiates an action potential, nominally -40 mV.
traveling waves: EEG oscillations that move across the cortex that may mediate large-scale coordination of brain networks and support connectivity.
transient: a single wave or sequence of regular waves, called a complex,
distinguishable from background EEG activity.
triphasic wave: a wave that contains three deflections from baseline.
unmyelinated axons: smaller-diameter axons without fatty insulation that
conduct more slowly than myelinated axons.
+ve: the source is the place at the other end of the neuron where current
leaves, and is represented by +ve.
-ve: a sink is where current enters the neuron. Positive sodium ion entry into
a neuron creates an active sink, represented by -ve.
ventral: toward the base of the skull or front of the body.
ventral striatum: the olfactory tubercle and nucleus accumbens.
ventral tegmental area: the midbrain structure that distributes dopaminergic axons
to the nucleus accumbens. Serotonin receptors on endorphin-releasing neurons in the hypothalamus may increase the
activity of dopaminergic reward pathways by inhibiting the release of GABA at receptors on cell bodies of the
ventral tegmental area neurons.
vigilance system: in Sterman’s model, a system that consists of both
specific brainstem nuclei (e.g., locus coeruleus and raphe nuclei) and their diffuse connections with the
thalamus and other subcortical structures, and the cortex. Several neurotransmitter systems mediate vigilance,
including cholinergic/glutamatergic (reticular formation), noradrenergic (locus coeruleus), and serotonergic
(raphe) neurons.
volume transmission: extrasynaptic neurotransmitter release from axonal
varicosities, dendrites, and terminal buttons into the extracellular space. Monoamines like norepinephrine and
serotonin are released outside the synaptic cleft.
waveform: the shape and form of an EEG signal.
white matter: the layer beneath the cortex that mainly consists of myelinated
axons.
TEST YOURSELF ON CLASSMARKER
Click on the ClassMarker logo below to take a 10-question exam over this entire unit.
REVIEW FLASHCARDS ON QUIZLET PLUS
Click on the Quizlet Plus logo to review our chapter flashcards.
Visit the BioSource Software Website
BioSource Software offers Functional Neuroanatomy, which reviews critical behavioral networks, and qEEG100, which provides extensive multiple-choice testing over the IQCB Blueprint.
Assignment
Now that you have completed this unit, how would you explain the relationship between local field potentials and the EEG? How does anatomy explain why the EEG is comprised of EPSPs and IPSPs instead of action potentials?
References
Advokat, C. D., & Comaty, J. E., & Julien, R. M. (2019). Julien's primer of drug action (15th ed.). Worth Publishers.
Altenmuller, E. O., & Gerloff, C. (1999). Psychophysiology and the EEG. In E. Niedermeyer & F. Lopes da Silva (Eds.), Electroencephalography: Basic principles, clinical applications, and related fields (4th ed.). Williams and Wilkins.
E. Niedermeyer & F. Lopes da Silva (Eds.) (1999). Electroencephalography: Basic principles, clinical applications, and related fields (4th ed.). Williams and Wilkins.
Andreassi, J. L. (2007). Psychophysiology: Human behavior and physiological
response (5th ed.). Lawrence Erlbaum and Associates, Inc.
Arnsten, A. F. (2006). Fundamentals of Attention-Deficit/Hyperactivity Disorder: Circuits and pathways. Journal
of Clinical Psychiatry, 67 (Suppl. 8), 7-12.
Babiloni, C., Babiloni, F., Carducci, F., Cincotti, F., Del Percio, C.,
Hallett, M., Moretti, D. V., Romani, G. L., & Rossini, P. M. High
resolution EEG of sensorimotor brain functions: Mapping ERPs or mu ERD?
In R. C. Reisin, M. R. Nuwer, M. Hallett, & C. Medina (Eds.). Advances
in Clinical Neurophysiology (Supplements to Clinical Neurophysiology
Vol. 54). Elsevier Science B. V.
Basile, L. F. H., Yacubian, J., Ferreira, B. L. C., Valim, A. C., & Gattaz, W. F. (2004). Topographic abnormality of slow cortical potentials in schizophrenia. Brazilian Journal of Medical and Biological Research, 37(1), 97-109. https://doi.org/10.1590/s0100-879x2004000100014
Bear, M. F., Connors, B. W., & Paradiso, M. A. (2020). Neuroscience:
Exploring the brain (4th ed.). Jones & Bartlett Learning.
Breedlove, S. M., & Watson, N. V. (2020). Behavioral neuroscience (9th ed.). Sinauer Associates, Inc.
Heinrich, H., Gevensleben, H., Freisleder, F. J., Moll, G. H., & Rothenberger, A. (2004). Training of slow cortical potentials in attention-deficit/hyperactivity disorder: Evidence for positive behavioral and neurophysiologic effects. Biological Psychiatry, 55(7), 772–775. https://doi.org/10.1016/j.biopsych.2003.11.013
Hugdahl, K. (1995). Psychophysiology: The mind-body perspective. Harvard University Press.
Kalat, J. W. (2019). Biological psychology (13th ed.). Cengage Learning.
Klein, S. B., & Thorne, B. M. (2007). Biological psychology. New York: Worth Publishers.
Kotchoubey, B., Blankenhorn, V., Fröscher, W., Strehl, U. & Birbaumer, N. (1997). Stability of cortical self-regulation in epilepsy patients. Neuroreport, 27(8)1867-1870. https://doi.org/10.1097/00001756-199705260-00015
Kotchoubey, B., Busch, S., Strehl, U. & Birbaumer, N. (1999). Changes in EEG power spectra during biofeedback of slow cortical potentials in epilepsy. Applied Psychophysiology and Biofeedback, 24(4) 213-233. https://doi.org/10.1023/a:1022226412991
Kotchoubey, B., Kubler. A., Strehl. U., Flor, H., & Birbaumer, N. (2002). Can humans perceive their brain states? Consciousness and Cognition, 11(1), 98-113. https://doi.org/10.1006/ccog.2001.0535
Kotchoubey, B., Schneider, D., Schleichert, H., Strehl, U., Uhlmann, C., Blankenhorn, V., Fröscher, W., & Birbaumer, N. (1996). Self-regulation of slow cortical potentials in epilepsy: A retrial with analysis of influencing factors. Epilepsy Research, 25(3), 269-276. https://doi.org/10.1016/s0920-1211(96)00082-4
Kotchoubey, B., Schneider, D., Uhlmann, C., Schleichert, H., & Birbaumer, N. (1997). Beyond habituation: Long-term repetition effects on visual event-related potentials in epileptic patients. Electroencephalographer, 103(4), 450-456. https://doi.org/10.1016/s0013-4694(97)00026-6
Kotchoubey, B., Strehl, U., Holzapfel, S., Blankenhorn, V., Fröscher. W., & Birbaumer, N. (1999). Negative potential shifts and the prediction of the outcome of neurofeedback therapy in epilepsy. Clinical Neurophysiology, 110(4), 683-686. https://doi.org/10.1016/s1388-2457(99)00005-x
Kotchoubey. B., Strehl, U., Holzapfel, S., Schneider. D., Blankenhorn, V., & Birbaumer, N. (1999). Control of cortical excitability in epilepsy. Adv Neurol, 81, 281-290.
Kotchoubey, B., Strehl, U., Uhlmann, C., Holzapfel, S., Konig, M., Fröscher. W., Blankenhorn, V., & Birbaumer, N. (2001). Modification of slow cortical potentials in patients with refractory epilepsy: A controlled outcome study. Epilepsia, 42(3), 406-416.
Kropotov, J. D. (2009). Quantitative EEG, event-related potentials and neurotherapy. Academic Press.
Munro, C. A., McCaul, M. E., Wong, D. F., Oswald, L. M., Zhou, Y., Brasic, J., Kuwabara, H., Kumar, A., Alexander, M., Ye, W., & Wand, G. S. (2006). Sex differences in striatal dopamine release in healthy adults. Biological
Psychiatry, 59(10), 966-974. https://doi.org/10.1016/j.biopsych.2006.01.008
Nash, J. M. (2011). The gift of mimicry. Your brain: A user's guide. Time.
Niedermeyer, E. (1999). Historical aspects. In E. Niedermeyer & F. Lopes da Silva (Eds.), Electroencephalography: Basic principles, clinical applications, and related fields (4th ed.). Williams and Wilkins.
E. Niedermeyer & F. Lopes da Silva (Eds.) (1999). Electroencephalography: Basic principles, clinical applications, and related fields (4th ed.). Williams and Wilkins.
Otto, D., & Reiter, L. (1984) Developmental changes in slow cortical potentials of young children with elevated body lead burden: Neurophysiological considerations. Annals of the New York Academy of Sciences, 425(1), 377-383. https://doi.org/10.1111/j.1749-6632.1984.tb23559.x
Pulvermüller, F., Mohr, B., Schleichert, H., & Veit, R., (2000). Operant conditioning of left-hemispheric slow cortical potentials and its effect on word processing. Biological Psychology, 53(2-3), 177-215. https://doi.org/10.1016/S0301-0511(00)00046-6
Raichle, M. E., & Gusnard, D. A. (2002). Appraising the brain's energy budget. PNAS, 99(16), 10237-10299. https://doi.org/www.pnas.orgcgidoi10.1073pnas.172399499
Schneider, F., Rockstroh, B., Heimann, H., Lutzenberger, W., Mattes, R., Elbert, T., Birbaumer, N, & Bartels, M. (1992). Self-regulation of slow cortical potentials in psychiatric patients: Schizophrenia. Biofeedback and Self-Regulation, 17(4), 277-292. https://doi.org/10.1007/bf01000051
M. S. Schwartz, & F. Andrasik (Eds.). (2003). Biofeedback: A
practitioner's guide (3rd ed.). The Guilford Press.
Sebel, P. S., Lang, E., Rampil, I. J., White, P. F., Cork, R., Jopling, M., Smith, N. T., Glass, P. S., & Manberg, P. (1997). A multicenter study of bispectral electroencephalogram analysis monitoring anesthetic effect. Anesthesia and Analgesia, 84(4), 891-899. https://doi.org/10.1097/00000539-199704000-00035
Siniatchkin, M., Hierundar, A., Kropp, P., Kuhnert, R., Gerber, W-D., & Stephani, U. (2000). Self-regulation of slow cortical potentials in children with migraine: An exploratory study. Applied Psychophysiology & Biofeedback, 25(1), 13-32. https://doi.org/10.1023/a:1009581321624
Speckmann, E.-J., & Elger, C. E. (1984). The neurophysiological basis of epileptic activity: A condensed overview. R. Degen & E. Niedermeyer (Eds.), Epilepsy, sleep, and sleep deprivation (pp.23-34). Elsevier. PMID: 1760082
Speckmann, E.-J., & Elger, C. E. (1999). Introduction to the neurophysiological basis of the EEG and DC potentials. In E. Niedermeyer & F. Lopes da Silva (Eds.), Electroencephalography: Basic principles, clinical applications, and related fields (4th ed.). Williams and Wilkins.
E. Niedermeyer & F. Lopes da Silva (Eds.) (1999). Electroencephalography: Basic principles, clinical applications, and related fields (4th ed.). Williams and Wilkins.
Stahl, S. M. (2008). Stahl’s essential psychopharmacology: Neuroscientific basis and practical
applications (3rd ed.). Cambridge University Press.
Steriade, M. (2005). Cellular substrates of brain rhythms. In E. Niedermeyer, & F. Lopes da Silva (Eds.). Electroencephalography:
Basic principles, clinical applications, and related fields (5th ed.). Lippincott Williams
& Wilkins.
Thompson, M., & Thompson, L. (2016). The biofeedback book: An
introduction to basic concepts in applied psychophysiology (2nd ed.). Association for Applied Psychophysiology and Biofeedback.
Thompson, M., & Thompson, L. (2009). Asperger’s syndrome intervention: Combining neurofeedback,
biofeedback, and metacognition. In T. H. Budzynski, H. K. Budzynski, J. R. Evans, & A. Abarbanel (Eds.). Introduction to quantitative EEG and neurofeedback (2nd ed.). Academic Press.
Warren, A. M., & McIlvane, W. J. (1998). Stimulus equivalence and the
N400 effect. Poster presented at the 1998 Annual Meeting of the Cognitive
Neuroscience Society in San Francisco, CA.
Wilson, J. (2003). Biological foundations of human behavior. Wadsworth/Thompson Learning.
Wilson, V. E., Thompson, M., Thompson, L., Thompson, J., Fallahpour, K., & Linden, M. K. (2011).
Introduction to biofeedback (Neurofeedback). In B. W. Strack, M. K. Linden, & V. S. Wilson (Eds.). Biofeedback
& neurofeedback applications in sport psychology. Association for Applied Psychophysiology and Biofeedback.
Winn, P. (2001). (Ed.), Dictionary of biological psychology. Routledge.





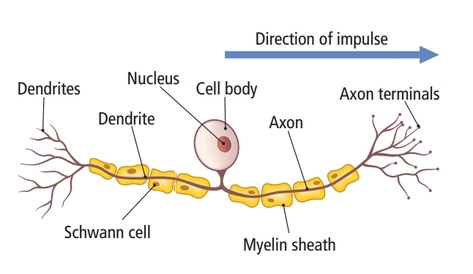
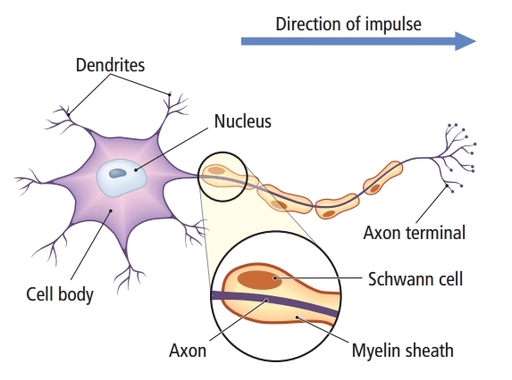
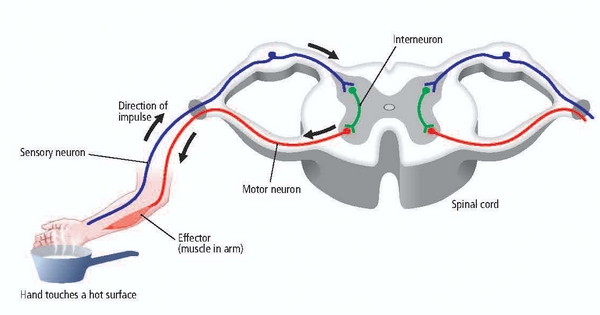
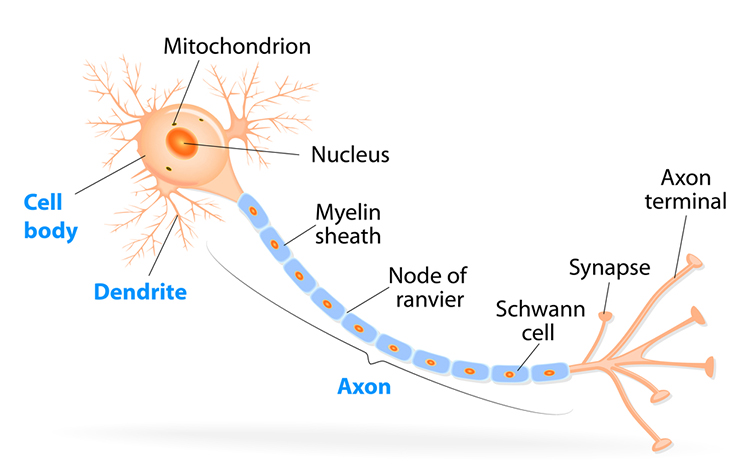
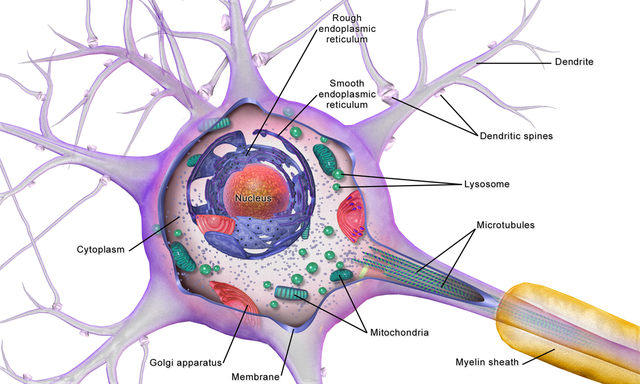
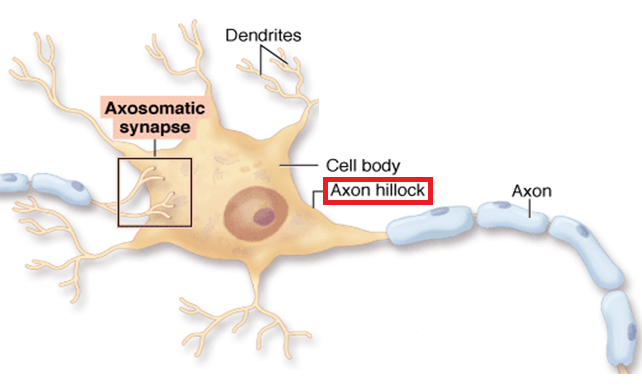
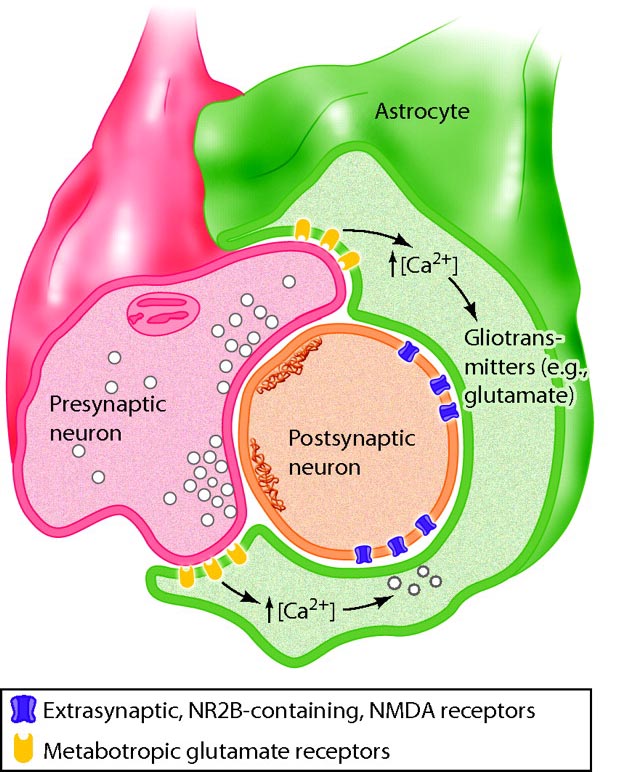
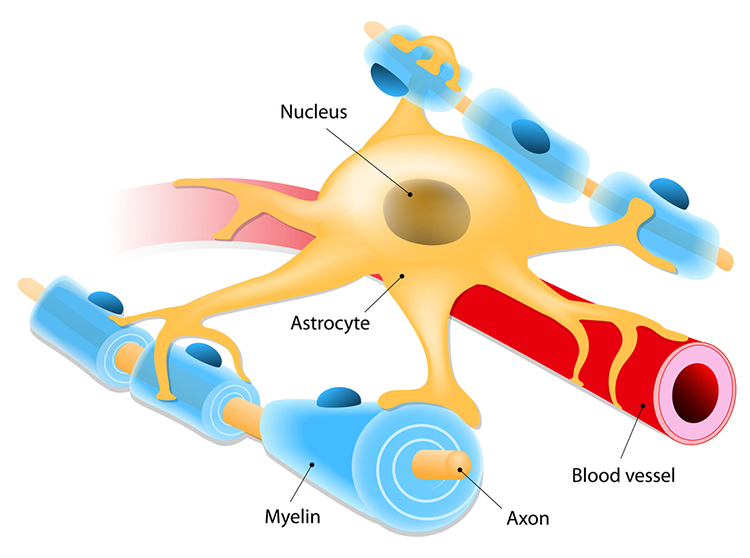
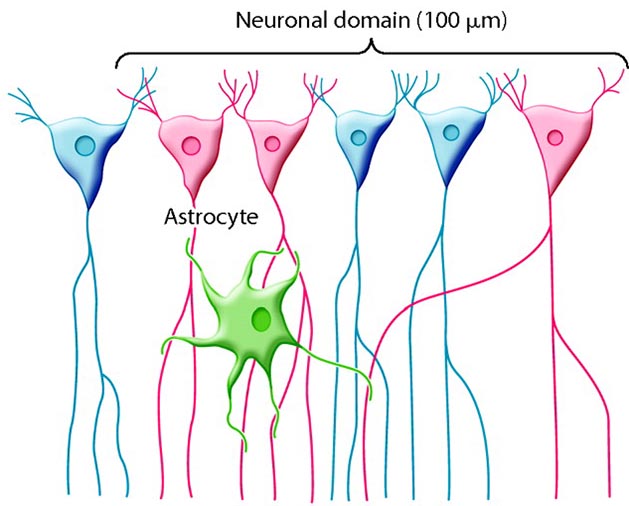
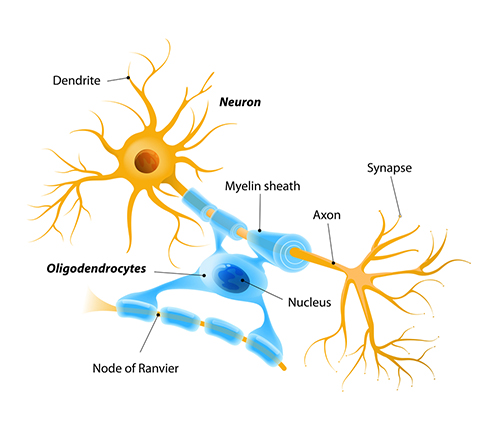
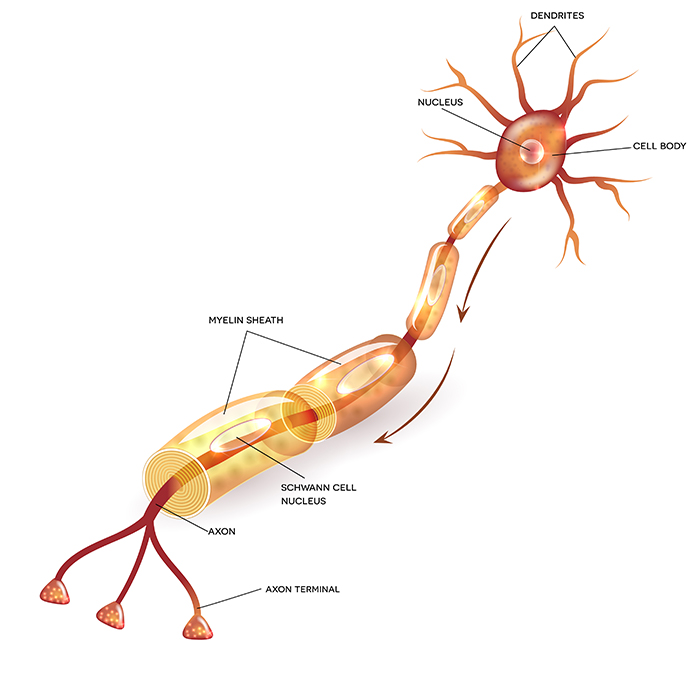
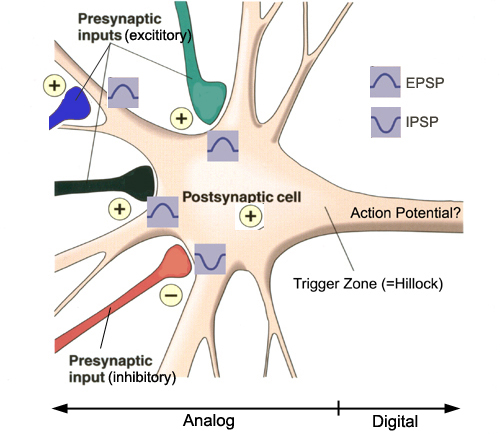
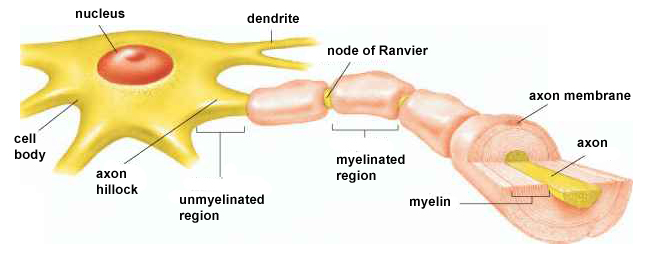
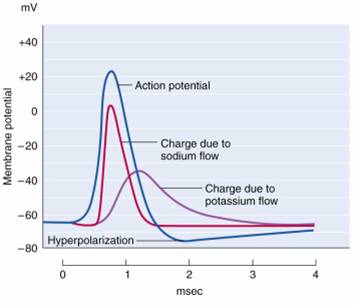
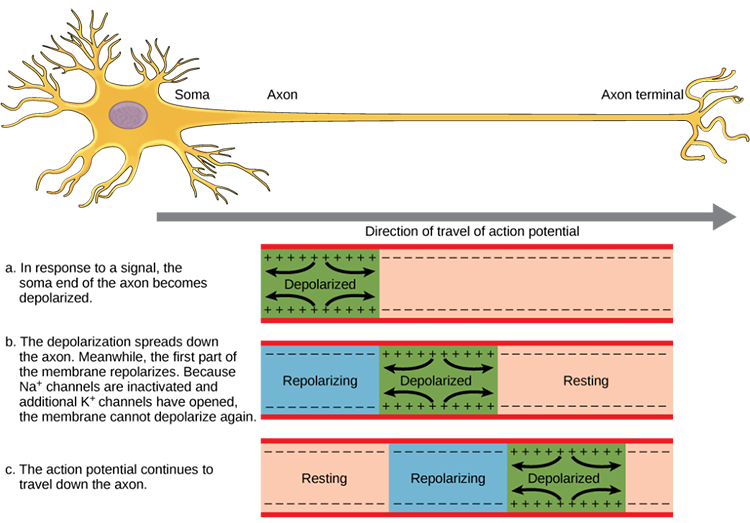
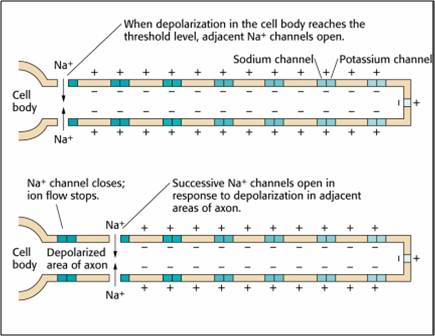
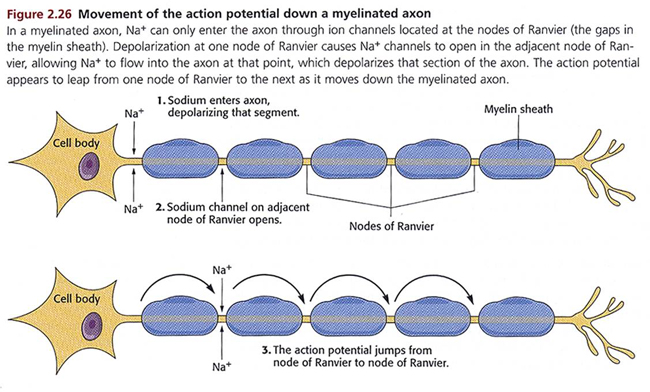
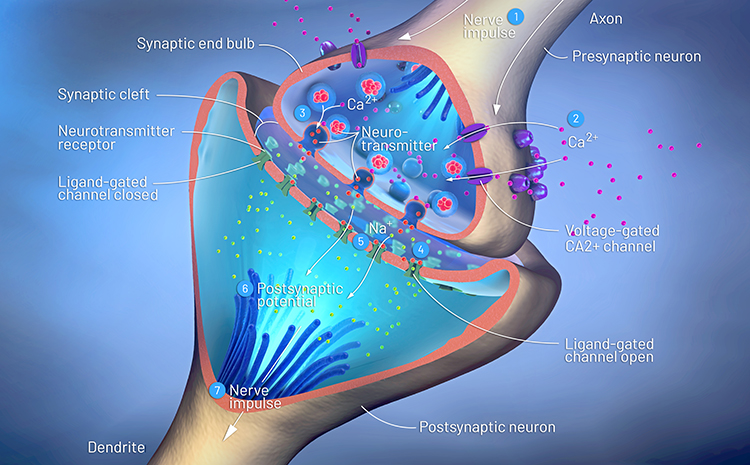

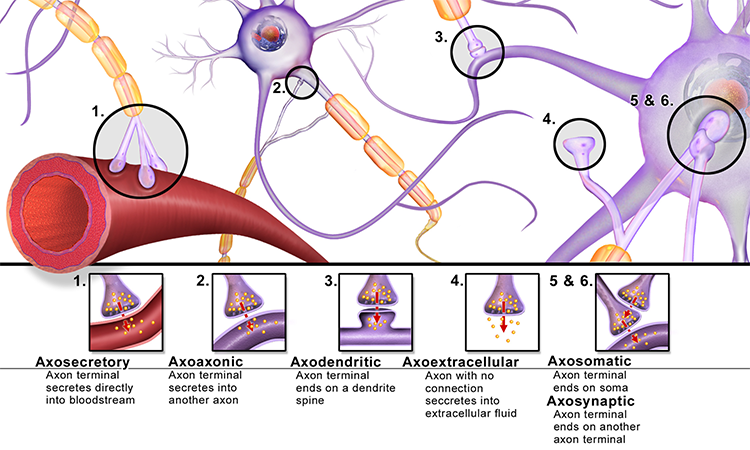

.jpg)