Neuroanatomy
What You Will Learn in This Chapter
This unit takes you on a comprehensive tour of neuroanatomy, from the ascending sensory pathways that bring information to the cortex to the thalamic and cortical generators that produce the EEG rhythms you will encounter in clinical practice. You will learn how the brain's major structures, including the frontal, parietal, temporal, and occipital lobes, contribute to cognition, emotion, and behavior.
You will also explore subcortical structures, Brodmann areas and their clinical relevance, and the connectivity, phase, and coherence concepts that underlie EEG network analysis. By the end of this chapter, you will understand how neuroanatomical knowledge informs neurofeedback assessment and training decisions.
BCIA Blueprint Coverage: This unit addresses II. Basic Neurophysiology and Neuroanatomy - B. Neuroanatomy.
The simplest way to divide the cortex is into the frontal and posterior regions. The frontal cortex (frontal lobe) specializes in action, encompassing cognition, emotion, autonomic control, movement, and speech. The posterior cortex (parietal, temporal, and occipital lobes) is concerned with perception and memory. Together with subcortical structures and the peripheral nervous system, these regions provide the hierarchically arranged feedback loops that allow us to interact with our environment to achieve goals successfully.
The prefrontal cortex (PFC), the cortex rostral to the motor association cortex, directs cognitive and emotional processes called perception-action cycles, which adapt (and preadapt) us to our environment. In essence, the PFC predicts and creates the future. Working in cooperation with networked brain structures, it marshals executive functions such as planning, attention, working memory, and decision-making to develop innovative actions in pursuit of future goals (Fuster, 2015).
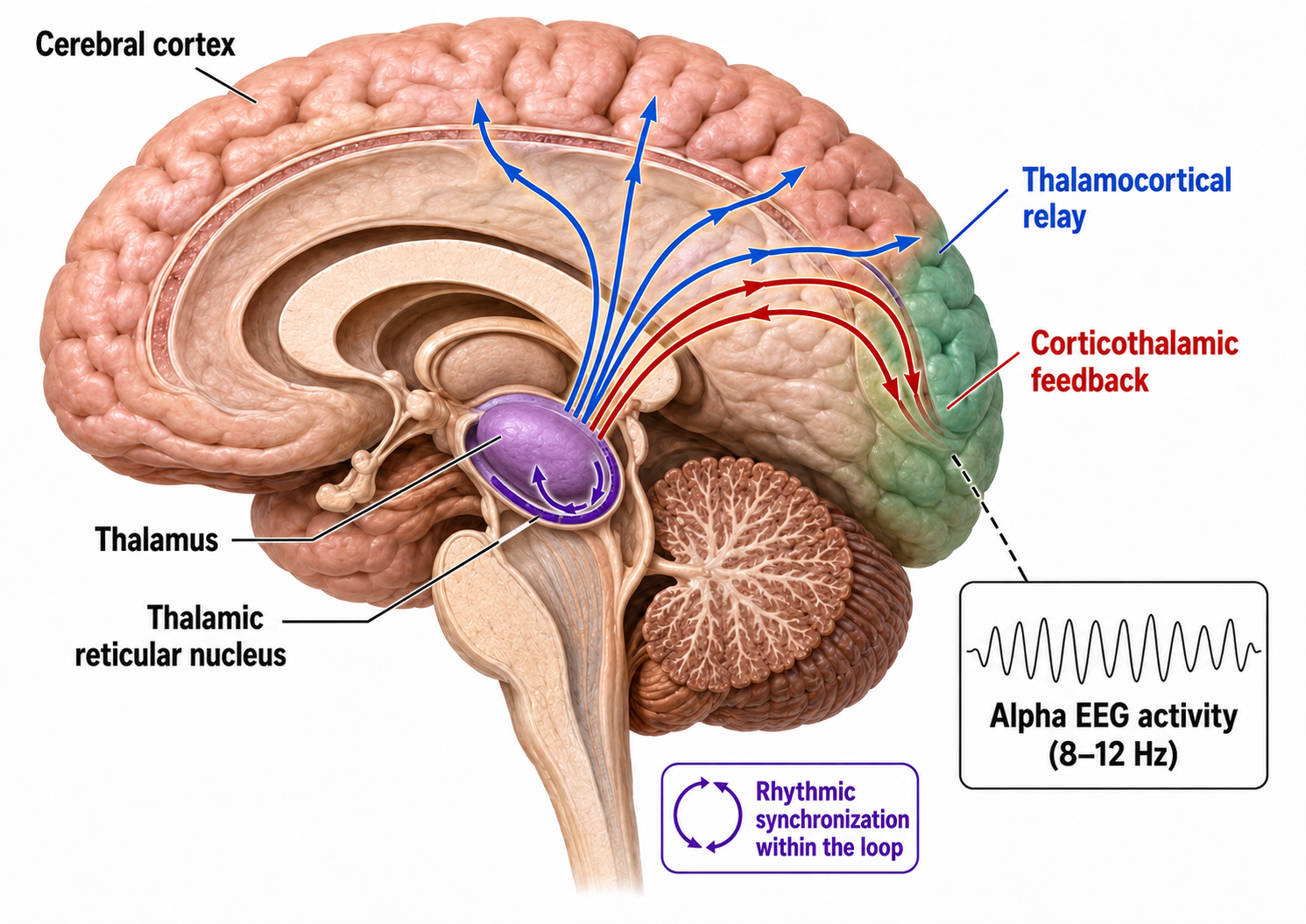
The nervous system uses bottom-up (feedforward) and top-down (feedback) processing to maintain homeostasis. This interconnectedness is best illustrated by the relationship between the thalamus and cortex: ascending thalamocortical neurons distribute sensory information to appropriate cortical and subcortical regions, while descending corticothalamic neurons convey instructions back to the thalamus. The nervous system generates EEG activity ranging from DC potentials to beta-gamma rhythms, using multiple generators that operate as what neuroscientist William Calvin called a "cerebral symphony."

Graphic © adike/Shutterstock.com.
BCIA Blueprint Coverage
This unit addresses II. Basic Neurophysiology and Neuroanatomy - B. Neuroanatomy.

This unit covers Basic Neuroanatomy of Ascending Sensory Pathways to the Cortex, Thalamic, Cortical, and Subcortical Generators of the EEG, General Cortical and Subcortical Anatomy, Major Functions of Cortical Lobes and Major Subcortical Structures and Brodmann Areas, and Overview of Connectivity, Phase, and Coherence Concepts Related to EEG Networks and Tracts.
Full-Length Lecture: Neuroanatomy
Basic Anatomy of Ascending Sensory Pathways to the Cortex
This section examines how sensory information travels from the body's receptors to the cortex, focusing on the visual, auditory, and somatosensory systems. Understanding these pathways is essential for biofeedback practitioners because sensory input directly shapes the EEG patterns you will assess and train. Sensory input produced by activities like reading a novel or listening to music can desynchronize cortical activity, resulting in lower-amplitude, higher-frequency EEG waveforms (Neumann, Strehl, & Birbaumer, 2003). Arousal and specific forms of cognitive activity may reduce alpha amplitude or eliminate it entirely, a phenomenon called alpha blocking, while increasing EEG power in the beta range (Andreassi, 2007).
The classical routes for EEG activation consist of ascending sensory pathways that distribute information to specialized thalamic nuclei, which then project the results of thalamic processing to the appropriate cortical regions. A hallmark of these pathways is their hierarchical structure, which preserves the spatial location from which signals arise, with specialized thalamic nuclei serving as critical relay points. The one exception is olfaction (smell), which bypasses the thalamus and projects directly to the primary olfactory cortex.
The old-school view held that these ascending pathways exercise purely bottom-up control of perception as feedforward circuits. This overlooks the fact that ten times more cortical efferent neurons target the sensory thalamus than thalamic afferent neurons project to the cortex. The new-school view recognizes extensive interconnections between the thalamus and cortex that permit a substantial degree of top-down cortical control over perception (Kandel et al., 2021). Subcortical areas, including the midbrain, thalamus, and cerebellum, also participate in ascending and descending transmission and processing of neural messages.
In summary, sensory pathways are not one-way streets but dynamic, bidirectional systems in which the cortex actively shapes the information it receives. We will now examine the visual, auditory, and somatosensory systems to illustrate these principles.
Visual System
Retinal ganglion cells, which comprise the optic nerves, ascend to the midline optic chiasm where the two optic nerves meet. At the chiasm, temporal axons (those toward the side of the head) continue on their own side's optic tract, while nasal axons cross over to join the opposite side's optic tract. Most optic tract axons project to the lateral geniculate nucleus (LGN) of the thalamus.
While the LGN relays visual information to the cortex, brainstem and cortical neurons actively modulate its activity. Brainstem neurons that mediate alertness and attention can adjust the LGN's response to visual input. The cortex can also exert top-down selection to increase attention to a salient region of the visual field at the expense of others (Bear, Connors, & Paradiso, 2020). This is clinically relevant because it demonstrates that even at the earliest relay station, perception is shaped by attention and arousal, the very processes biofeedback practitioners aim to train.
LGN neurons form the optic radiations and project to the primary visual cortex (V1) in the occipital lobe's cortical layer IV. A minority of retinal ganglion cell axons target the dorsal midbrain superior colliculus, which directs visual gaze and selective attention to visual objects (Breedlove & Watson, 2023). The cortex contains many specialist regions for processing visual properties like color, shape, location, motion, and orientation, organized into two major streams originating in V1.
The dorsal stream projects from V1 to the parietal lobe, helping us localize objects and guide movements. The adjacent motor association cortex contains neurons with both visual and motor properties, called mirror neurons, whose networks may play a role in learning actions by observing others, understanding intentions, and empathy. The lower ventral stream projects to inferior temporal and frontal areas and allows us to identify objects and faces.
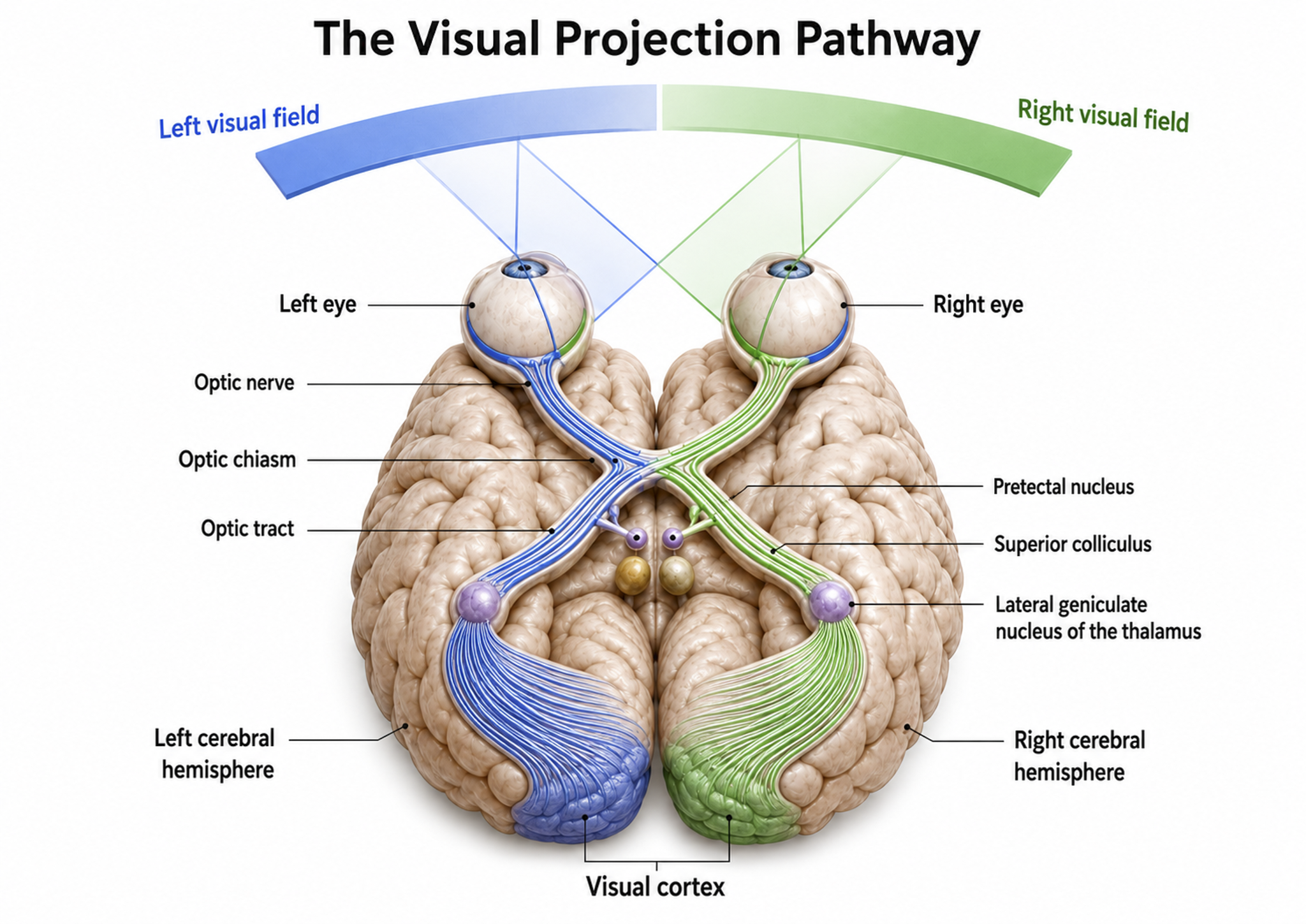
Visual pathway graphic adapted from © Alila Medical Media/Shutterstock.com.
Auditory System
The cochlea's inner hair cells within the organ of Corti send approximately 30,000 auditory fibers to several destinations: the superior olivary nuclei of the pons and the midbrain inferior colliculi, and the medial geniculate nucleus of the thalamus. The superior olivary nuclei process binaural (two-ear) information to localize sound, while all ascending auditory neurons ultimately innervate the inferior colliculi, some via intermediate relays.
The inferior colliculi integrate information about spatial localization and multiple sensory modalities, including somatosensory information. They project to the thalamus' medial geniculate nucleus (MGN), which in turn projects to several cortical auditory areas using two separate pathways. The MGN mainly relays frequency, amplitude, and binaural information to the auditory cortex in the temporal lobe.
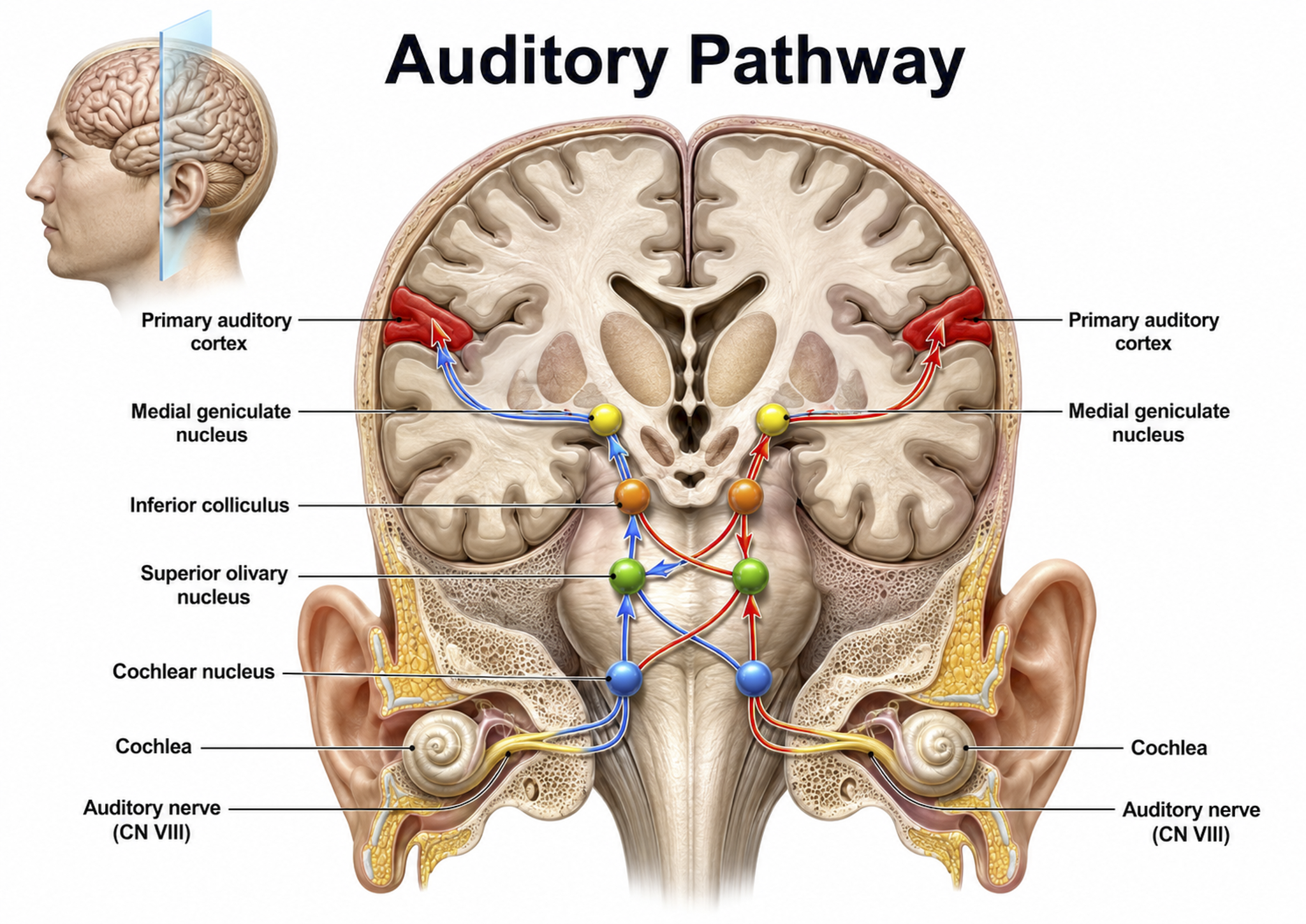
Auditory pathway graphic © medicalstocks/Shutterstock.com.
The auditory cortex processes auditory information within dorsal and ventral streams, paralleling the visual system's organization. The dorsal stream, extending to the parietal lobe, helps us spatially localize sounds. The lower ventral stream, projecting to the temporal lobe, analyzes sound components, possibly including speech sounds (Breedlove & Watson, 2023). As with the visual pathways, the auditory system involves extensive feedback: brainstem neurons innervate outer hair cells that adjust basilar membrane sensitivity to specific frequencies, and auditory cortex axons innervate both the inferior colliculi and MGN to exercise top-down control (Bear, Connors, & Paradiso, 2020).
Somatosensory System
The somatosensory system employs specialized receptors to perceive itch, pain, temperature, and touch. For touch, the axon of a unipolar neuron enters the dorsal horn of the spinal cord and synapses with a dorsal column neuron in the medulla. Axons from this neuron decussate (cross the midline) and innervate the thalamus' ventral posterior nucleus (VPN), which in turn distributes this information to the primary somatosensory cortex (S1).
While each hemisphere's S1 maps touch information from the opposite side of the body, the secondary somatosensory cortex (S2) maps both sides. The maps are overlaid so that the left and right arms are represented in the same region of the body surface map (Breedlove & Watson, 2023). This bilateral representation in S2 may help explain why somatosensory processing can produce widely distributed EEG changes that clinicians observe during assessment.
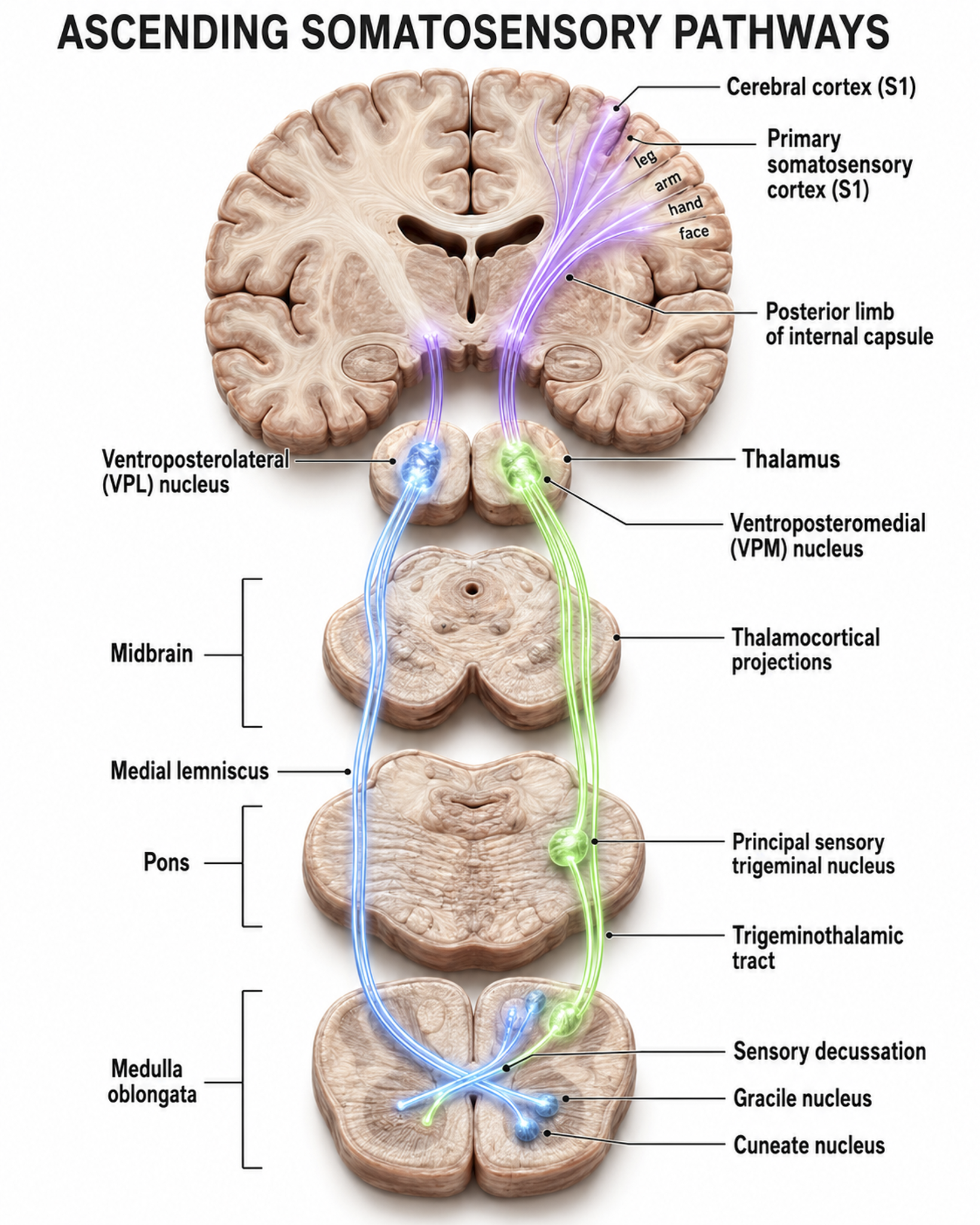
As with the visual and auditory systems, the ascending somatosensory pathways do far more than relay information. These networks process and alter sensory information at each successive synapse, and the cortex exercises top-down control over neurons in the dorsal column and VPN to dynamically adjust cortical inputs (Bear, Connors, & Paradiso, 2020). The recurring theme across all three sensory systems is bidirectional communication: the cortex is not a passive receiver but an active participant that shapes the sensory data it processes.
Clinical Application
Imagine Maria, a graduate student who closes her eyes during an assessment and produces a strong posterior alpha rhythm. The moment you ask her to solve a mental arithmetic problem, her alpha amplitude drops and beta activity rises. This is alpha blocking, the desynchronization that sensory input and focused cognition produce (Andreassi, 2007). Recognizing that ordinary attention reshapes the EEG helps you set realistic baselines and interpret changes during training rather than mistaking them for pathology.
Key Takeaways
Ascending sensory pathways relay information through specialized thalamic nuclei to the appropriate cortical regions, preserving the spatial location from which signals arise. Olfaction is the exception, bypassing the thalamus to reach the primary olfactory cortex directly. These pathways are not one-way streets, because roughly ten times more cortical neurons project back to the sensory thalamus than the other way, giving the cortex substantial top-down control. The visual, auditory, and somatosensory systems each split into a dorsal stream for locating stimuli and a ventral stream for identifying them. Sensory input and arousal desynchronize the EEG, lowering alpha and raising beta, a change clinicians see as alpha blocking.
Check Your Understanding
- Which sensory modality bypasses the thalamus, and where does it project instead?
- What does the roughly ten-to-one ratio of corticothalamic to thalamocortical projections tell us about how perception is controlled?
- Trace the visual pathway from the retinal ganglion cells to the primary visual cortex, naming the thalamic relay.
- How does sensory input change the EEG, and what is the phenomenon called when alpha is reduced or eliminated?
Thalamic, Cortical, and Subcortical Generators of the EEG
This section explores how the brain generates the EEG rhythms central to neurofeedback practice. We will cover thalamic pacemakers, cortical resonant loops, and subcortical modulators, then survey each major EEG frequency band. Understanding these generators helps clinicians interpret why certain rhythms appear at specific scalp locations and what they signify about brain function.
Thalamic Generators
Andersen and Andersson (1968) advanced the facultative pacemaker theory, proposing that thalamic neurons activate both cortical neurons and thalamic inhibitory interneurons via recurrent collaterals. While these thalamocortical neurons only excite a limited number of cortical neurons, the inhibitory interneurons suppress a large pool of thalamocortical relay neurons. When the inhibition ends after about one-tenth of a second, the relay neurons experience rebound excitation, a synchronized depolarization that excites both cortical neurons and another round of thalamic inhibitory interneurons, initiating a new cycle that produces rhythmic EEG activity (Fisch, 1999).
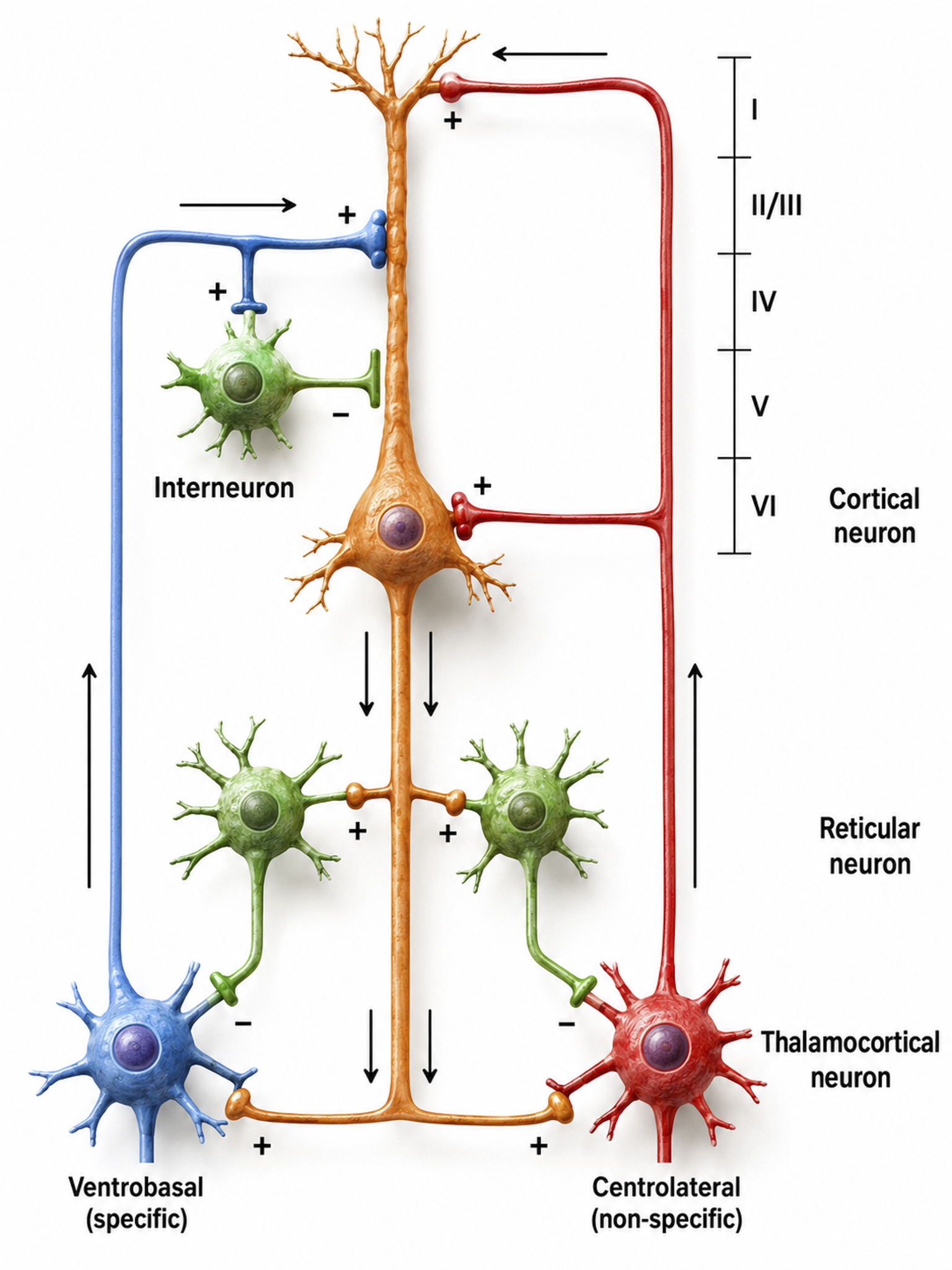
Thalamocortical circuit diagram depicting specific/sensory and non-specific intralaminar thalamocortical systems. Graphic adapted from Zachary Barry and featured in Wikipedia's article Recurrent ThalamoCortical Resonance.
The networking of excitatory and inhibitory thalamic neurons imposes a group rhythm on its members, which is then transmitted to cortical macrocolumns by thalamocortical neurons (Bear, Connors, & Paradiso, 2020). The nucleus reticularis of the thalamus may function as a pacemaker by releasing the inhibitory transmitter GABA at synapses with thalamocortical neurons. When this inhibition ends, these neurons depolarize cortical neurons and thalamic inhibitory interneurons through burst discharges.
As discussed in the Neurophysiology unit, oscillatory activity may involve an interaction between thalamocortical relay neurons (TCR), nucleus reticularis neurons (RE), and interneurons, mediated by diverse neurotransmitters including acetylcholine and GABA. The thalamus is the dominant pacemaker for rhythmic EEG activity, including theta (4-7 Hz), alpha (8-12 Hz), and SMR (12-15 Hz) (Amzica & Lopes da Silva, 2018). For biofeedback practitioners, this means that many of the rhythms you train have their origin not in the cortex beneath the electrode, but in deeper thalamic circuits.
Cortical Generators
The cerebral cortex (gray matter) consists of neuronal cell bodies, glial cells, and blood vessels, while white matter lies beneath it, composed of myelinated nerves, nonmyelinated fibers, and glial cells. The EEG mainly originates from pyramidal neurons in layers III, V, and VI of the gray matter, the six-layered outer portion of the cerebrum that contains neuronal cell bodies. Cortical thickness ranges from about 1 to 4.5 mm and averages roughly 2.5 mm.
Why do pyramidal neurons dominate the scalp EEG? Their long apical dendrites run parallel to one another and perpendicular to the cortical surface, so that synchronous synaptic currents along these dendrites create aligned electrical dipoles whose fields add together rather than cancel, a configuration known as an open field (Kirschstein & Köhling, 2009). Neurons whose dendrites radiate symmetrically in all directions, such as many stellate and thalamocortical cells, instead form a closed field in which opposing currents largely cancel, so they contribute little to the scalp signal (Buzsáki, Anastassiou, & Koch, 2012). Because each dipole is tiny, thousands of neighboring pyramidal neurons must depolarize or hyperpolarize together for their summed field to survive volume conduction through the meninges, skull, and scalp and reach a recording electrode (Buzsáki et al., 2012).
Key Concept
The scalp EEG is dominated by cortical pyramidal neurons because their long apical dendrites run parallel to one another and perpendicular to the cortical surface. When these neurons are active together, their aligned dipoles form an open field whose electrical fields summate rather than cancel. Neurons whose dendrites radiate symmetrically instead form a closed field in which opposing currents largely cancel, so they contribute little to the scalp signal. Because each dipole is tiny, thousands of neighboring pyramidal neurons must depolarize or hyperpolarize together, and a reliable scalp EEG requires at least six square centimeters of synchronized cortex. What summates is their synchronized postsynaptic activity, not their rate of action potentials.

Pyramidal neuron graphic © Juan Gaertner/Shutterstock.com.
Vertical cortical macrocolumns, on the order of a millimeter across, contain hundreds of thousands of pyramidal neurons together with supporting stellate and basket cells (Thompson & Thompson, 2015). Each cortical pyramidal neuron may receive on the order of 5,000 to 30,000 synapses. These macrocolumns are positioned side by side and perpendicular to the cortical surface, and since neighboring macrocolumns often receive the same afferent messages, they tend to fire together and generate a potential detectable from the scalp. A reliable scalp EEG requires a minimum of 6 cm² of synchronized cortex (Dyro, 1989).
Although thalamic pacemakers generate EEG rhythms, resonant loops between cortical macrocolumns represent another source (Traub et al., 1989). Over 97 percent of the brain's conversations are cortical-to-cortical, primarily within the same hemisphere (Thompson & Thompson, 2015). A resonant loop develops when macrocolumns that share afferent input fire synchronously to generate an electrical potential. The distance between participating macrocolumns is one determinant of EEG frequency: the closer the macrocolumns, the higher the frequency they can generate (Lubar, 1997).
There are three types of resonant loops driven by afferent input or thalamic pacemakers. Local loops couple neighboring macrocolumns and may generate frequencies above 30 Hz in the high-beta and gamma ranges. Regional loops couple macrocolumns separated by several centimeters and may produce alpha and beta rhythms. Global loops couple macrocolumns as distant as 7 cm (for example, between the frontal and parietal lobes) and may create delta and theta rhythms. The graphic below is conceptual, and should not be taken literally with respect to distance or anatomical location.
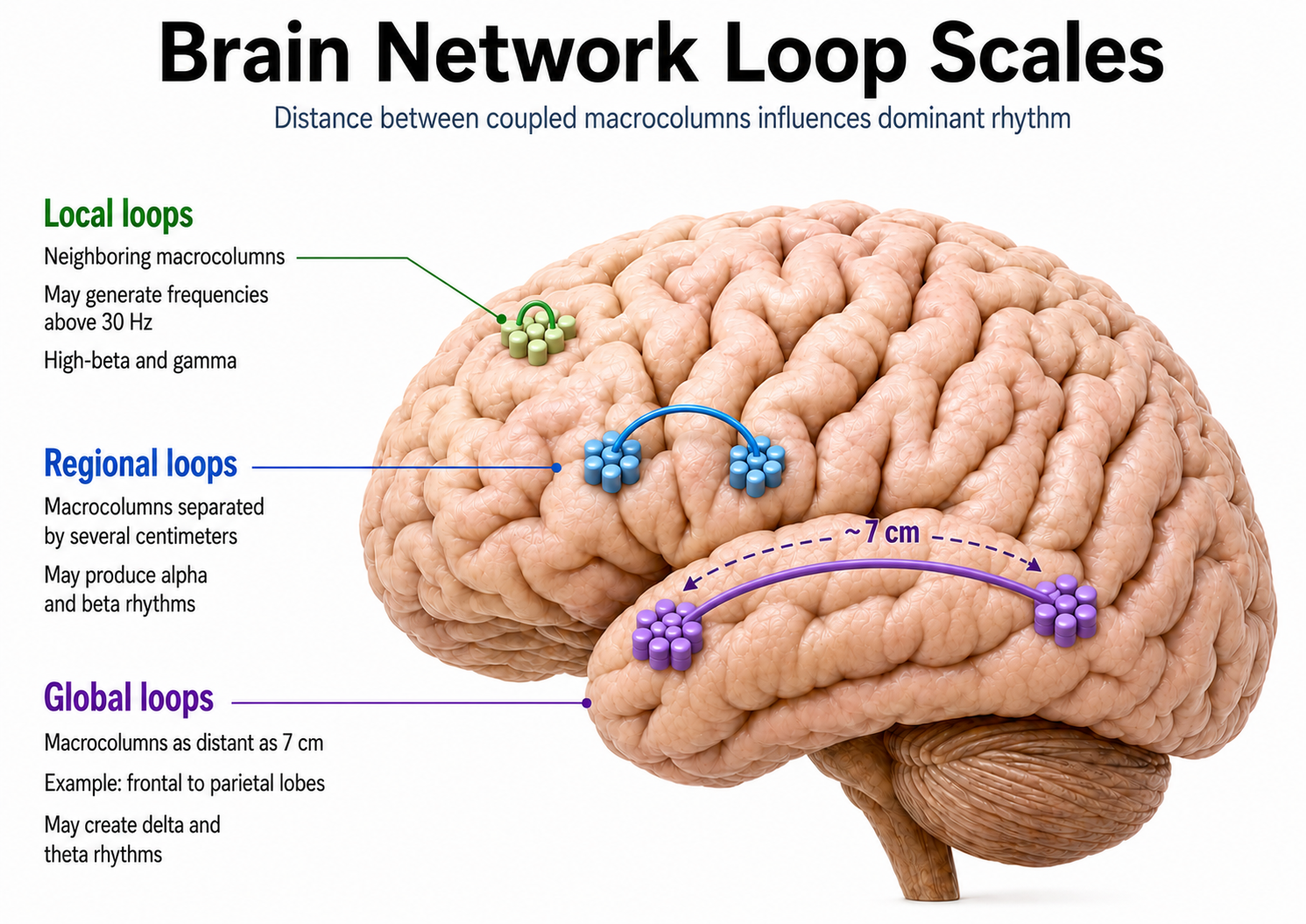
While only 3 percent of linkages are thalamocortical, they greatly influence the EEG by subcortically connecting distant cortical regions and producing most synchronous activity (Steriade, 2005).
Lubar (1997) proposed a violin analogy: the thalamic pacemakers firing at varying frequencies are the strings, and the resonant loops introducing different time delays are the instrument's resonant cavity.
Spindling is a synaptically generated oscillation in a circuit that includes the reticular nuclei (Steriade, 2005). The video of alpha spindling © John S. Anderson.
Different spindle frequencies result from corresponding durations of thalamocortical neuron hyperpolarization. Longer hyperpolarizations associated with EEG synchronized states produce 7-Hz or lower-frequency spindles, while relatively short hyperpolarizations result in 14-Hz spindles (Steriade, 2005). The electrical potentials generated by the thalamus can volume conduct near the speed of light through cerebrospinal fluid (CSF), brain tissue, the skull, and the scalp, so that nearly identical waveforms can simultaneously appear at distant sites (Fisch, 1999; Thompson & Thompson, 2015). This is an important consideration when interpreting EEG topographic maps. Activity at a given electrode may not originate from the cortex directly beneath it.
The Locus Coeruleus Inhibits Thalamic Alpha Generators
When we are inattentive, thalamic pacemakers generate the alpha rhythm. When we need to focus attention, we activate the brainstem noradrenergic locus coeruleus, a small but powerful nucleus roughly 15 mm long. The increased release of norepinephrine by this structure focuses attention and abolishes alpha oscillations by suppressing thalamic alpha generators. This may be an underlying mechanism of the phenomenon of alpha blocking.
Although researchers cannot noninvasively monitor locus coeruleus activity in human participants, it correlates with pupil dilation. In human studies, the greater the alpha blocking response and pupil dilation, the better the performance on demanding attention tasks (Dahl et al., 2020; Dahl et al., 2022). Critically, the alpha rhythm is not a cause but a sign that incoming stimulation is too weak to overcome inhibition by the reticular nucleus. EEG activity is not causal; it reflects network activity that has already occurred.
Additional Subcortical Generators
Several subcortical systems desynchronize or disrupt brain rhythms through ascending projections. These include the basal forebrain, reticular formation, locus coeruleus, and raphe systems. These neurons receive information from most sensory systems and cortical regions, and they desynchronize the EEG both directly through synapses on cortical neurons and indirectly through innervation of thalamic pacemakers. Desynchronization shifts pyramidal neurons from burst firing to more continuous single-spike generation (Fisch, 1999).
The cholinergic basal forebrain, located in the ventral frontal lobe and anterior hypothalamus, influences cerebral blood flow and cognitive activity. The basal forebrain comprises several structures, including the basal nucleus of Meynert, medial septal nuclei, substantia innominata, nucleus accumbens, ventral pallidum, and olfactory tubercle. Of these, the basal nucleus of Meynert and the medial septal nuclei are the principal sources of the neurotransmitter acetylcholine.
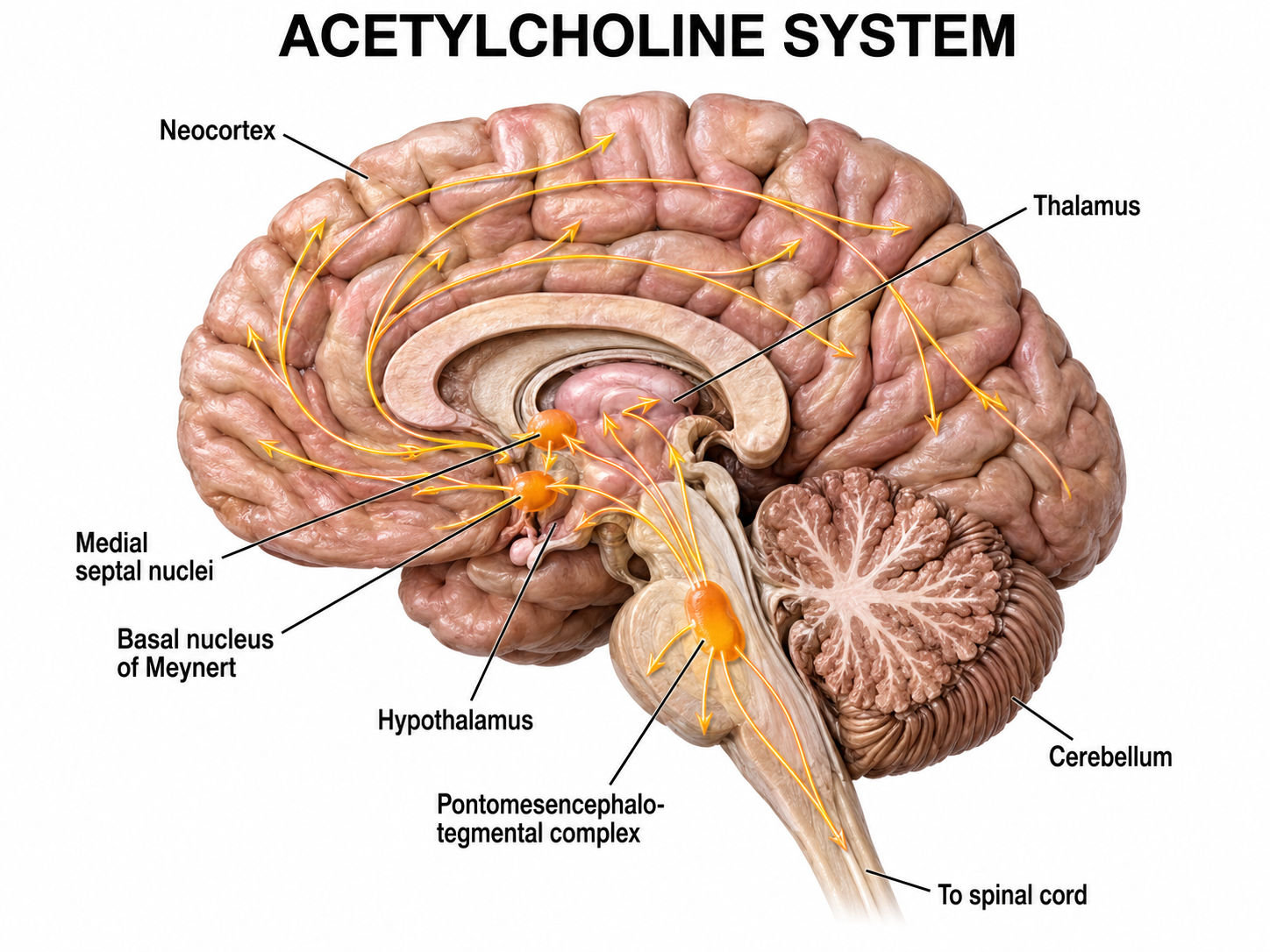
Graphic adapted from © Vasilisa Tsoy/Shutterstock.
The reticular activating system (RAS) includes a network of roughly 90 nuclei within the central brainstem, extending from the lower medulla to the upper midbrain, that activates the brain to promote attention, consciousness, and wakefulness. This network receives input from ascending sensory tracts (auditory, olfactory, somatosensory, and visual systems) and projects both to the thalamus and diffusely to the cortex, including pathways that bypass the thalamus entirely.
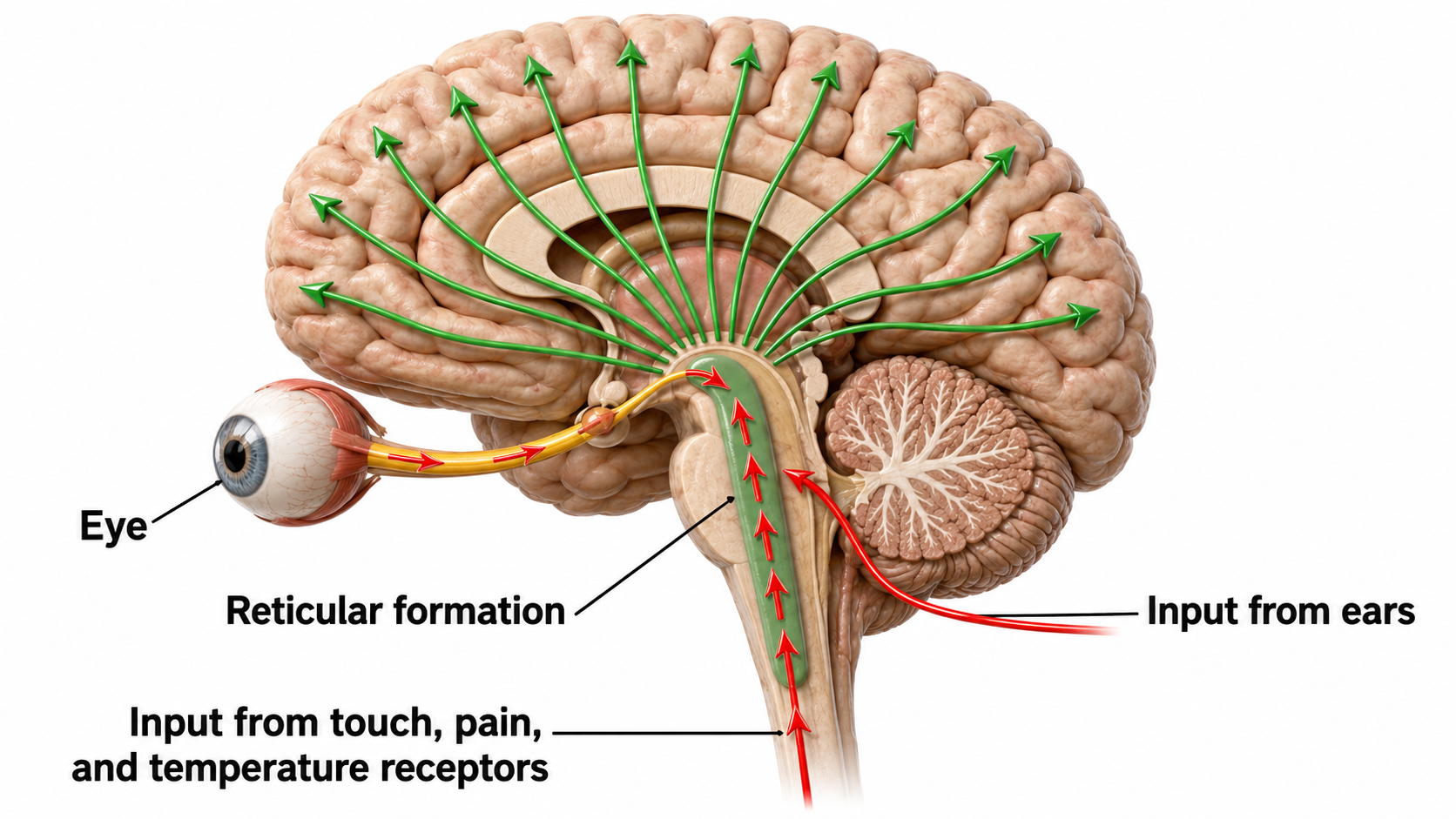
Reticular formation graphic redrawn by minaanandag on Fiverr.com.
The noradrenergic brainstem locus coeruleus system projects to the thalamus, limbic system, and cerebral cortex and contributes to wakefulness and vigilance for salient stimuli. The neurotransmitter norepinephrine (also called noradrenaline) is synthesized chiefly by neurons of the locus coeruleus, a small, pigmented nucleus situated in the dorsal pons along the lateral floor of the fourth ventricle. Its axons project diffusely throughout the neuraxis, reaching the cerebral cortex, hippocampus, amygdala, thalamus, and cerebellum, along with descending fibers to the spinal cord. Through these widespread connections, the locus coeruleus regulates arousal, selective attention, the sleep-wake cycle, and the physiological response to stress.
Within these neurons, norepinephrine is produced through a sequential enzymatic pathway. Tyrosine hydroxylase converts the amino acid tyrosine to L-DOPA, aromatic L-amino acid decarboxylase then yields dopamine, and dopamine β-hydroxylase completes the final step inside synaptic vesicles. Although the locus coeruleus supplies most of the forebrain's norepinephrine, additional noradrenergic populations in the lateral tegmentum and medulla contribute to autonomic and brainstem circuits.
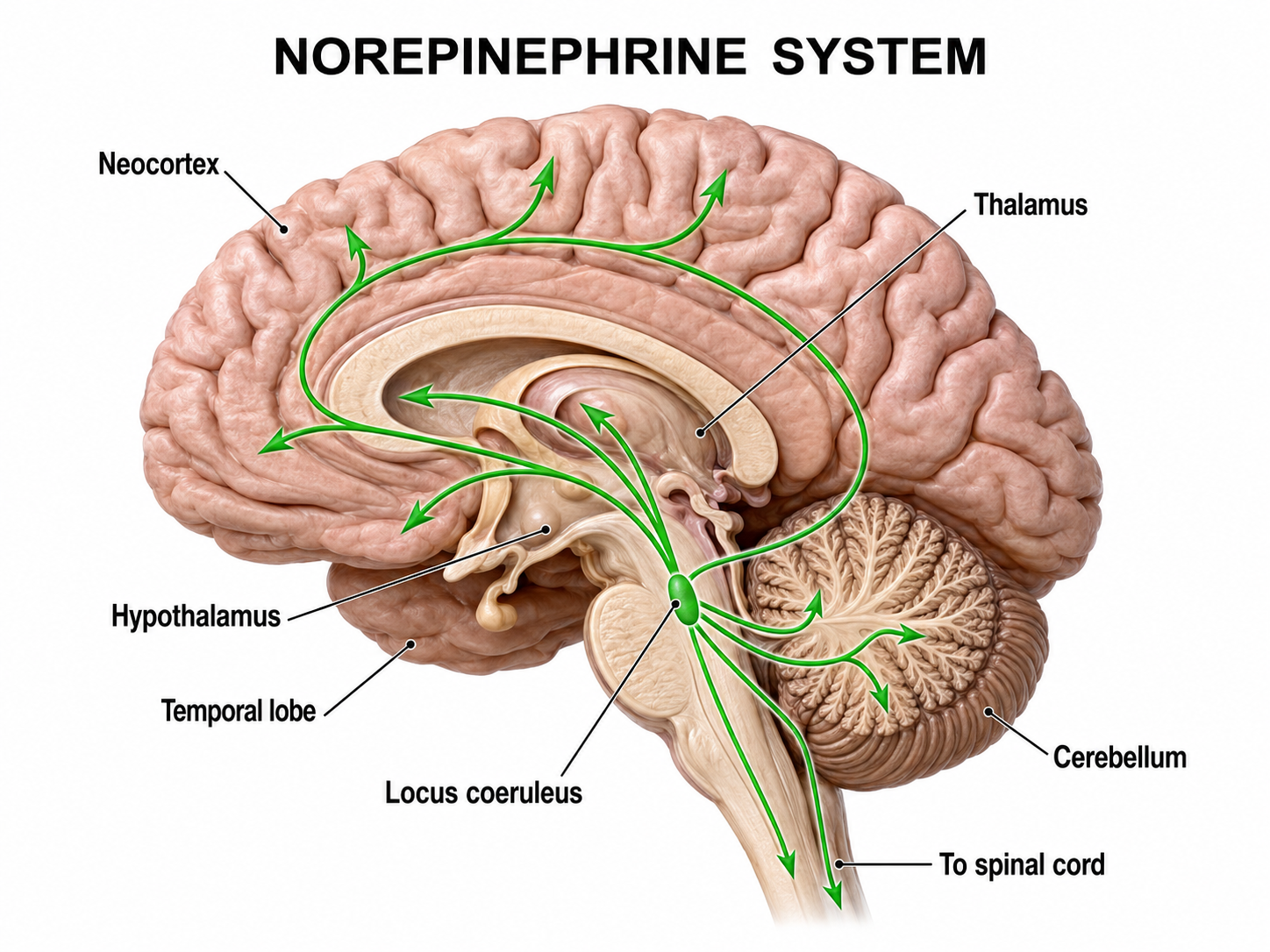
Graphic adapted from © Vasilisa Tsoy/Shutterstock.
Finally, the serotonergic raphe system is a midline network of cell bodies within the brainstem and midbrain that may influence alertness and vigilance through reciprocal connections with the suprachiasmatic nucleus of the hypothalamus (Monti & Jantos, 2008). Together, these subcortical systems determine the brain's overall state of arousal, a critical variable in any biofeedback session.
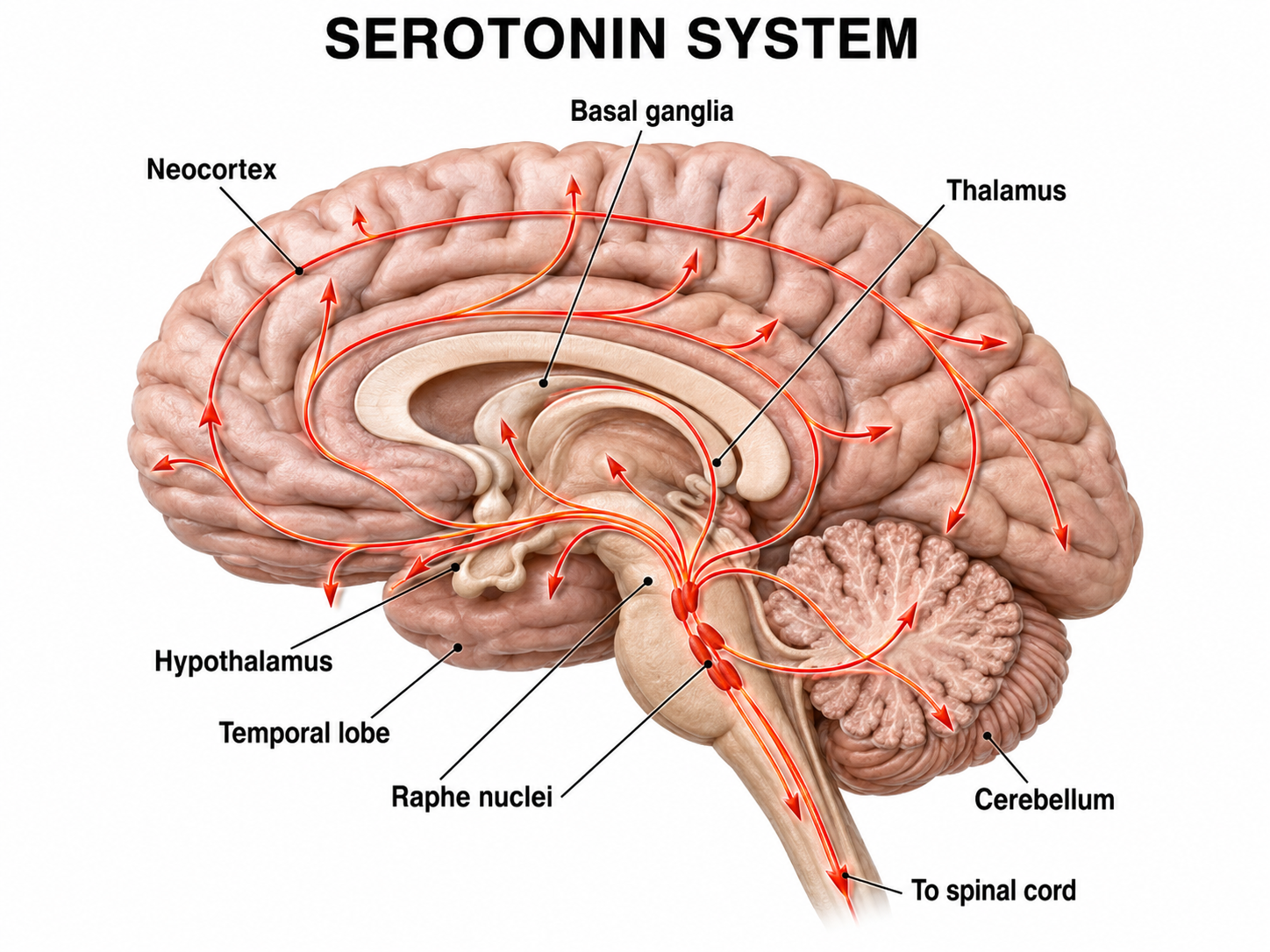
Graphic adapted from © Vasilisa Tsoy/Shutterstock.
Cortical and Subcortical Generators of Specific EEG Rhythms
Slow Cortical Potentials (0-1 Hz)
A slow cortical potential (SCP) is a gradual, sustained shift in the cortex’s baseline voltage that lasts from roughly 300 milliseconds to several seconds and reflects changes in the excitability of large populations of cortical neurons (Birbaumer et al., 1990). Slow cortical potentials (SCPs) have been identified in cortical neurons, the thalamus, and glial cells. Cortical neurons in layers II to VI generate slow oscillations even when the thalamus is removed or when cortical tissue is studied in vitro (in an artificial environment) or in vivo (within a living organism). Thalamic reticular neurons exhibit similar slow spontaneous oscillations when studied in vitro, and synchronized intracortical oscillations may depend on a corticothalamic network that targets these thalamic neurons.
The prevailing account attributes surface-negative SCPs to the synchronous depolarization of pyramidal apical dendrites, with a contribution from glial cells rather than a purely glial origin. Glia contribute by buffering extracellular potassium and by coupling to one another through gap junctions, which are direct electrical connections between glial cells. Glial cells communicate among themselves and with neurons, and their slow oscillations may influence the timing of neuronal firing through their control of potassium ion outflow (Steriade, 2005). These slow oscillations appear to organize the generation of other brain rhythms.
"The concept of a unified corticothalamic network that generates diverse types of brain rhythms grouped by the cortical slow oscillation (Steriade, 2001a,b) is supported by EEG studies in humans" (Mölle et al., 2002). Caton (1875) reported feeble currents of varying direction from electrodes placed on the cortical surface and observed that the surface becomes relatively negative during functional activity. Caton recorded galvanometer deflections rather than calibrated voltages, so his one-page report contains no microvolt figures; the numerical gradients sometimes attributed to him do not appear in it. Underlying "tone" or valence factors determine the firing characteristics of neurons within a network: when SCPs are more positive, reduced cortical neuron firing occurs due to hyperpolarization; when SCPs are more negative, increased firing occurs due to depolarization.
The following 19-channel BioTrace+ /NeXus-32 display of 0.1-1 Hz SCP activity © John S. Anderson.
Perspective on Fast Cortical Potentials
EEG "bands" are somewhat arbitrary ranges of frequencies that have evolved from observation and usage. The following BioTrace+ /NeXus-32 video of raw and spectral EEG displays © John S. Anderson. Frequency is plotted along the horizontal axis, and amplitude is shown on the vertical axis.
While frequency band labels are helpful descriptors, they can also be misleading. Classification of an EEG rhythm is based on context (measurement conditions and EEG activity during the specific epoch), frequency, and waveform morphology. Note, however, that for some purposes it is useful to define individual-specific frequency bands that depart somewhat from the classical definitions. For example, although the individual alpha peak typically sits near 10 Hz, some individuals peak nearer 8 or 9 Hz and others nearer 12 Hz, and this shift can affect the settings selected for neurofeedback (Tarasi & Romei, 2024).
Cutting Edge
Classical band boundaries are useful but not absolute. Because individual alpha frequency varies from person to person, some clinicians define individualized frequency bands that depart from the classical definitions, and this shift can change the settings chosen for neurofeedback (Tarasi & Romei, 2024).
The process of up-training or down-training signal amplitude in one or more of the EEG bands using an EEG is called EEG biofeedback or neurofeedback. Minimum EEG voltages of 20-30 μV are seen in children and adults (Krauss et al., 2011).
Brainwaves Reflect Behavior
The ratio of slow (theta) to faster (beta) brainwaves indicates how alert you are; this is the theta/beta ratio. A higher ratio suggests greater drowsiness or inattention, while a lower ratio reflects focused alertness. This metric is one of the most widely used in neurofeedback assessment.
In the next section, we will examine delta, theta, rhythmic slow-wave, alpha, mu, synchronous "alpha," SMR, beta, high or fast beta, and gamma activity.
Local Versus Global Decision-making
The short time windows of fast oscillators facilitate local integration and decision-making, primarily because of the limitations of axon conduction delays. In contrast, the long time windows of slow oscillators can involve many neurons in large or distant brain areas, favoring complex, global decisions. This principle has direct clinical implications: when you observe a client's EEG dominated by slow activity, it may reflect an over-reliance on global, less differentiated processing at the expense of focused, efficient local integration.
Delta (1-4 Hz)
There are two delta rhythms: a slow oscillation under 1 Hz and a traditional 1-4 Hz oscillation. The slow 0.3-0.4 Hz oscillation originates in the neocortex and persists when the thalamus is removed, while thalamocortical neurons generate the 1-4 Hz oscillations observed during human stage-3 sleep. Slow neocortical oscillations may synchronize the thalamic delta rhythm (Steriade, 2005).
Delta activity is generated by cortical neurons when other connections do not activate them and is found predominantly in frontal areas. Delta is associated with sleep and infancy, and during stage 3 (N3) sleep it accompanies the replenishment of astrocyte glycogen stores. Clinicians observe delta in clients diagnosed with ADHD, brain tumors, learning disorders, and traumatic brain injury (TBI). Rhythmic high-amplitude delta is particularly associated with TBI when localized, while diffuse delta may appear in ADHD and learning disorders.
Delta waves are the main EEG activity during infancy, reflecting the immature brain's low-frequency cortical activity. As neural networks develop and mature, faster rhythms like alpha and beta replace delta dominance in waking states. In adults, waking delta activity is minimal but may transiently appear during drowsiness or relaxation.
Normal Amplitudes
Delta should not be present in significant amounts in the awake adult EEG. "Apparent" delta is usually an eye movement artifact, although some delta activity probably occurs in the normal waking adult EEG. In clinical practice, delta bands are inhibited or down-trained but rarely rewarded, though delta desynchronization can be rewarded.
The following 19-channel BioTrace+ /NeXus-32 display of eyes-open 1-4 Hz activity from a 10-year-old male © John S. Anderson.
Theta (4-7 Hz)
Researchers define the boundaries of EEG frequency bands somewhat differently; theta is variously set at 3-8, 4-7, or 4-8 Hz. This curriculum uses 4-7 Hz throughout, which is also the boundary used by the glossary entries for the septohippocampal system and the entorhinal cortex. Where a cited study or a training display uses a wider band, the study's own boundary is retained and named.
The mechanisms that generate the theta rhythm are poorly understood, and theta's functions differ depending on location and source. Amzica and Lopes da Silva (2018) consider the classic septal/diagonal band pacemaker model incomplete. Hippocampal interneurons exercise top-down control over the hypothetical medial septum pacemaker, and the hypothalamic supramammillary nucleus, with extensive connections to the brainstem, diencephalon, and medial septum, may also pace and modulate hippocampal theta. A non-cholinergic theta source has also been found within the entorhinal cortex of the hippocampus.
Theta is associated with creativity, global synchronization, memory formation, and recall. Increased theta amplitudes correspond with hypo-perfusion (reduced blood flow) and decreased glucose metabolism. Excessive frontal theta is linked with depression, daydreaming, distractibility, and inattention. Theta/beta (T/B) ratios are developmentally mediated, so any cutoff has to be age-referenced. In Monastra et al. (1999), the control mean fell from roughly 3.0 at ages 6-11 to roughly 1.5 at ages 21-30, and the ADHD thresholds fell in parallel (see the table below). A ratio of 3.0 is therefore unremarkable in a 6-to-11-year-old and markedly elevated in a young adult.
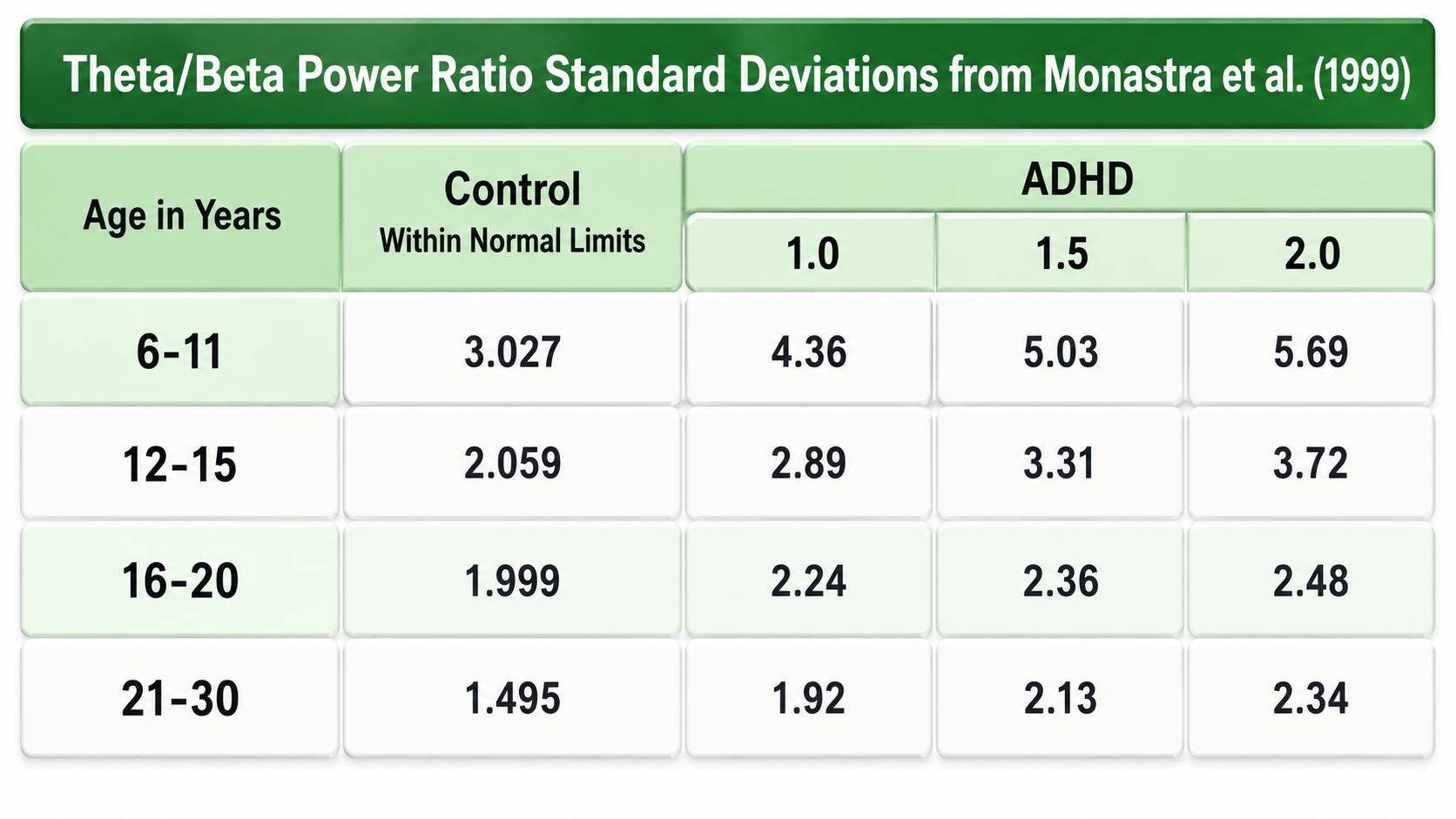
Normal Amplitudes
Theta voltage is age-related in the awake EEG, diminishing from age 8 to 30 with minimal amounts over age 30. A typical 6-7 Hz rhythm in the frontal midline (FCz) is associated with mental activity such as problem-solving and a wide variety of other functions. This rhythm appears to be limbic in origin and is higher in amplitude and more synchronous when processing feedback that an error has occurred. The 4-Hz rhythm is associated with childhood pleasurable experiences and memory searches in adults.
Rhythmic Slow Wave (RSW or Theta)
Clinicians inhibit frontal theta to remediate symptoms and reward posterior RSW in alpha/theta training for addictions, global synchronization, optimal performance, and PTSD. RSW is generally not increased frontally. Clinicians may also train for increases or decreases in phase synchrony. RSW is mainly seen in the frontal-midline (FCz) when awake with eyes open and is generated by the limbic system and thalamus. Depending on location, RSW may represent slowed alpha as thalamic output decreases in frequency.
The following 19-channel BioTrace+ /NeXus-32 display of eyes-open 4-8 Hz activity from a 10-year-old boy © John S. Anderson.
Alpha (8-12 Hz)
The 8-12-Hz alpha rhythm differs from spindle waves in both its source and the activity during which it is observed. Alpha 1 (low alpha) ranges from 8-10 Hz, and alpha 2 (high alpha) from 10-12 Hz (Thompson & Thompson, 2015). Some authors extend the alpha band to 13 Hz; this unit uses 8-12 Hz throughout. Alpha rhythms depend on the interaction between rhythmic burst firing by a subset of thalamocortical (TC) neurons linked by gap junctions and rhythmic inhibition by widely distributed reticular nucleus neurons (Hughes & Crunelli, 2005). The alpha rhythm is maintained and propagated by cortical networks (Amzica & Lopes da Silva, 2018).
Connectome graphic © Image Source Trading Ltd/Shutterstock.
Researchers have correlated the alpha rhythm with relaxed wakefulness, though there are age- and function-related differences. Spindle waves, in contrast, originate in the thalamus and occur during unconsciousness and stage-2 sleep (Steriade, 2005). Alpha is the dominant rhythm in adults and is located posteriorly.
The 8-10 Hz range is associated with ADHD, daydreaming, fogginess, OCD, and TBI, while frontal asymmetry with excess left frontal alpha is associated with depression. The 10-12 Hz range is seen with inner calm (calm and alert) and meditation. Clinicians train alpha amplitude and phase synchrony up or down depending on location and presenting symptoms.
Posterior Dominant Rhythm (PDR)
The posterior dominant rhythm (PDR) is the posterior alpha rhythm, first visible at about 4 months of age with a frequency of around 4 Hz. Between 3 and 5 years, this rhythm is approximately 8 Hz with amplitudes as high as 100 μV. From 6 to 15 years, it reaches 9 Hz by age 7 and 10 Hz by ages 10-15, with a mean amplitude of 50-60 μV. Girls show a statistically faster maturation of posterior alpha frequency than boys.
From 13 to 21 years, the mean alpha frequency is 10 Hz, and amplitudes decline throughout this period. Faster alpha frequencies are associated with higher IQ and better memory performance. This developmental trajectory illustrates why age-appropriate normative comparisons are essential in clinical qEEG assessment.
The following 19-channel BioTrace+ /NeXus-32 display of the response of the posterior dominant rhythm to eyes opening and closing © John S. Anderson.
Normal Amplitudes
The typical adult alpha peak frequency ranges from 9.5-10.5 Hz, and alpha below 8 Hz is considered abnormal. Alpha frequency and amplitude decline throughout adulthood and into old age. Most adult amplitudes fall below 60 μV: about 66% of adults show 20-60 μV, 28% show less than 20 μV, and 6% show greater than 60 μV (Schomer & Lopes da Silva, 2011).
Higher alpha amplitudes are observed over the non-dominant (right) hemisphere, a normal pattern known as alpha asymmetry. Most studies show no effect of handedness. Asymmetry is generally no more than 20 μV or 20% of the greater of the two amplitudes (Amzica & Lopes da Silva, 2018). Clinicians should be familiar with these normative values, since departures from them often guide neurofeedback protocol selection.
Causes of Excessive Alpha Amplitudes
Sleep deprivation or metabolic exhaustion can result in high amplitude and slowing of the peak frequency, along with persistent alpha during an eyes-open condition. Meditation practices can cause increased amplitudes and slowing, a faster alpha response to an eyes-closed condition, and persistent alpha in an eyes-open condition.
Marijuana use and abuse can also cause increased amplitudes and slowing, along with persistent alpha in an eyes-open condition, depending on the type of marijuana. These effects can persist for many years following abstinence. Clinicians should consider substance use history when interpreting alpha findings.
The following 19-channel BioTrace+ /NeXus-32 display of eyes-closed 8-12 Hz activity from a 13-year-old girl © John S. Anderson.
Mu Rhythm (7-11 Hz)
While the 7-11-Hz mu rhythm usually overlaps with the alpha range, it is arc-shaped on top and pointed on the bottom of an analog EEG trace (i.e., a wicket rhythm) that differentiates it from the alpha waveform. The mu rhythm is also identified by its location over the sensorimotor strip. It can be recorded at C3 and C4 in a minority of subjects and appears when the person is awake and at rest. Actual, visualized, or observed movement suppresses it (Thompson & Thompson, 2015). Mu rhythms appear to index sensorimotor cortex activity, and mu suppression has been linked to the mirror neuron system of the premotor and inferior parietal cortex.
These mirror neuron circuits may play a critical role in imitation learning and our ability to understand the actions of others. Mu rhythms facilitate the conversion of visual and auditory input into integrated skill-building functions, and attenuation of the mu rhythm appears to be associated with the activation of this function (Pineda, n.d.). The mu rhythm is highlighted below.
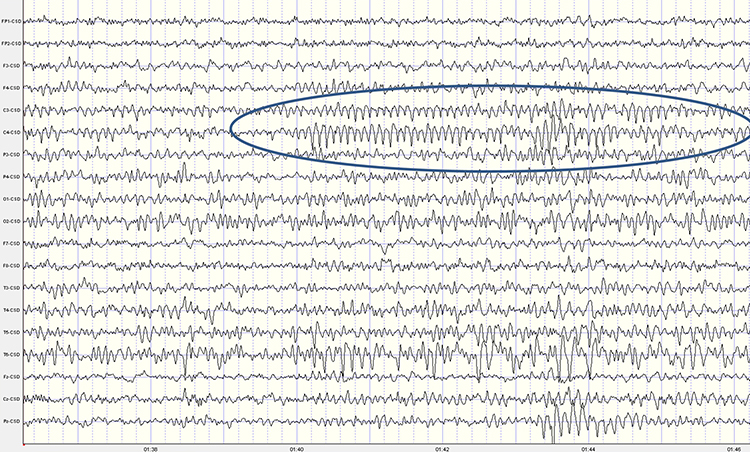
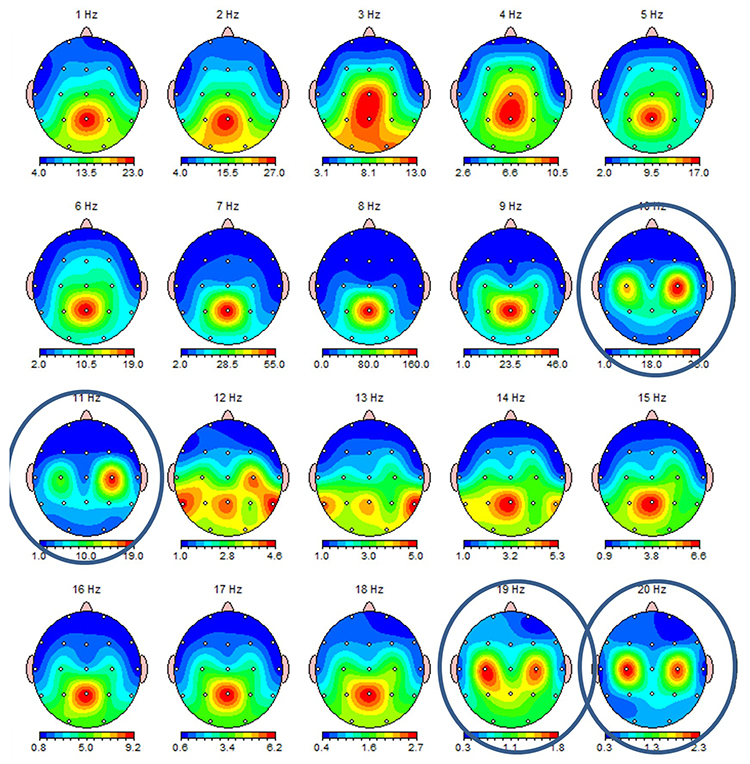
This second example of the mu rhythm shows a classic 10-11 Hz and 19-20 Hz "Owl Eye" presentation. The mu rhythm has an arciform, comb-like shape that produces two spectral peaks in a nearly harmonic relationship: a fundamental near 10 Hz and a beta harmonic near 20 Hz (Tiihonen, Kajola, & Hari, 1989). In the single-hertz topographic maps below, the mu fundamental near 10-11 Hz and its beta component near 19-20 Hz each appear as a pair of left and right central hot spots that look like an owl's eyes on the head map, which is the source of the name. The term is informal field usage rather than a published descriptor.
Synchronous "Alpha"
Various sensory systems (auditory, somatosensory, and visual) produce localized and semi-independent "alpha" activity. However, synchronous, distributed alpha integrates perception and facilitates action across broader cortical networks. Synchronous "alpha" appears to block localized alpha-like patterns such as mu and the posterior rhythm in favor of more broadly distributed network integration during tasks requiring global processing.
Sensorimotor Rhythm (12-15 Hz)
The sensorimotor rhythm (SMR), also called beta 1, is located on the sensorimotor strip (C3, Cz, C4). SMR amplitude increases when the motor circuitry is idle; it increases with stillness and decreases with movement. Deficient SMR may be observed in movement-spectrum complaints like hyperactivity and tics. SMR appears as sleep spindles during stage-2 sleep and is associated with reduced blood perfusion and glucose metabolism in the brain as it transitions to deeper sleep stages.
Clinicians typically reward increased SMR amplitude to calm hyperactivity and during theta/beta ratio training. For practitioners working with athletes or military personnel, SMR training can promote the calm, focused stillness essential for precision tasks like marksmanship or surgical procedures.
The following 19-channel BioTrace+ /NeXus-32 display of 12-15 Hz activity © John S. Anderson.
Beta (over 12 Hz)
Beta consists of rhythmic activity between 12-38 Hz in four ranges: beta 1 (12-15 Hz), beta 2 (15-18 Hz), beta 3 (18-25 Hz), and beta 4 (25-38 Hz). However, writers may define beta subranges somewhat differently.
Beta shows a gradient of activity from occipital to frontal lobes, with more beta occurring frontally where it is associated with focus, analysis, and relaxed thinking (Thompson & Thompson, 2015). Excessive beta is observed in anxiety, depression (reversed asymmetry), insomnia, OCD, and sleep disorders, while deficient beta is seen in ADHD, cognitive decline, and learning disorders.
Since beta overlaps with the EMG range, clinicians must be careful when up-training this rhythm and should use an EMG inhibit, (a high-beta EEG proxy for EMG), to avoid inadvertently reinforcing muscle artifact. Beta is generated by the brainstem and cortex and is associated with hyper-perfusion and increased glucose metabolism.
Normal 16-20+ Hz Beta Amplitudes
Beta amplitudes are minimal in children up to 12 years, with a significant increase in beta amplitude and organization between 12 and 30 years. Beta is commonly seen in nearly all adults with amplitudes of 20 μV or less. Interhemispheric amplitude asymmetries exceeding 35% are abnormal.
The following 19-channel BioTrace+ /NeXus-32 display of 13-21 Hz activity © John S. Anderson.
Fast or High Beta Rhythms (20-35 Hz)
Fast 20-35-Hz oscillations are generated by activation of the mesencephalic reticular formation. Thalamocortical, rostral thalamic intralaminar, and cortical neurons spontaneously oscillate in this range. This activity is primarily seen in the frontal lobes and is associated with hyper-perfusion and increased glucose metabolism, though persistent excessive activity can lead to metabolic exhaustion.
Fast beta may be associated with peak performance and cognitive processing, particularly with specificity and precision in information processing. However, excessive high beta is associated with alcoholism, anxiety, OCD, rumination, and worry. Clinicians often inhibit high beta activity but rarely reward it. In clinical practice, persistent high beta at frontal sites, particularly if it shows a spindling appearance, is one of the most common EEG signatures of anxiety disorders.
The following 19-channel BioTrace+ /NeXus-32 display of eyes-closed approximately 25 Hz fast beta activity © John S. Anderson.
Gamma Rhythms (28-80 Hz)
Because this unit ends beta at 38 Hz and begins gamma at 28 Hz, the two bands overlap between 28 and 38 Hz; clinical electroencephalography more often ends beta near 30 Hz. Any scalp band edge above roughly 30 Hz is also vulnerable to EMG contamination. Amzica and Lopes da Silva (2018) concluded that gamma oscillations might speed information distribution and processing. Gamma bursts occur during problem-solving, and the absence of gamma is associated with cognitive deficits and learning disorders. Gamma synchrony is related to cognitive processing and contributes specificity and precision to information processing. Gamma is theorized to serve as a "binding rhythm" that integrates sensory inputs into unified perception and consciousness.
The following 19-channel BioTrace+ /NeXus-32 display of eyes-open 36-44 Hz activity in a 10-year-old boy © John S. Anderson.
Gamma rhythms are linked with SCPs. The following BioTrace+ /NeXus-32 display of SCP and gamma activity © John S. Anderson.
In summary, the brain generates EEG rhythms through an interplay of thalamic pacemakers, cortical resonant loops, and subcortical modulators. Each frequency band reflects distinct physiological processes and clinical states, from delta's association with sleep and injury to gamma's role in cognitive binding. Understanding these generators equips clinicians to make informed decisions about which rhythms to train and why.
Clinical Application
Suppose you are training a client to increase sensorimotor rhythm at Cz and notice nearly identical activity appearing at neighboring sites. Remember that the rhythms you train often originate in deeper thalamic circuits rather than in the cortex directly beneath the electrode, and that electrical potentials volume conduct rapidly through tissue, skull, and scalp. As a result, activity recorded at one electrode may not arise from the cortex immediately below it. Keeping the thalamic origin of these rhythms and the reality of volume conduction in mind will make your interpretation of topographic maps more accurate.
Key Takeaways
The thalamus is the dominant pacemaker for rhythmic EEG activity, including theta, alpha, and the sensorimotor rhythm. Cortical rhythms also arise from synchronized pyramidal neurons in an open-field arrangement and from resonant loops, whose local, regional, and global forms generate progressively lower frequencies as the distance between macrocolumns increases. Subcortical systems, including the basal forebrain, reticular activating system, locus coeruleus, and raphe nuclei, desynchronize and modulate these rhythms to set the brain's level of arousal. Each frequency band reflects distinct states, from delta in sleep and injury to gamma in cognitive binding, with alpha marking relaxed wakefulness and beta marking focus or, in excess, anxiety. Because potentials volume conduct through tissue and skull, activity at a given electrode may not originate from the cortex directly beneath it.
Check Your Understanding
- According to the facultative pacemaker theory, what produces rhythmic EEG activity after thalamic inhibition ends?
- Why do pyramidal neurons, rather than stellate neurons, dominate the scalp EEG?
- Distinguish local, regional, and global resonant loops, and explain how the distance between macrocolumns relates to EEG frequency.
- What happens to the alpha rhythm when the locus coeruleus increases norepinephrine release, and what noninvasive measure correlates with this activity?
- Name the EEG band associated with each of the following: slow-wave sleep, relaxed wakefulness with eyes closed, and calm motor stillness on the sensorimotor strip.
General Cortical and Subcortical Anatomy
This section provides an overview of the nervous system's major divisions, from the central and peripheral nervous systems to the brain's subdivisions, protective structures, and vascular supply. These anatomical foundations are essential for understanding where EEG signals originate and how they are altered before reaching scalp electrodes.
Major Divisions
The human nervous system consists of the central nervous system and peripheral nervous system. The central nervous system (CNS) consists of the brain, spinal cord, and retina. Central nervous system graphic © SciePro/Shutterstock.
The 3-pound brain consists of approximately 86 billion neurons (Azevedo et al., 2009; Voytek, 2013). Graphic © Jasada Sabai/Shutterstock.com.
The cylindrical spinal cord extends from the medulla (brainstem) to the lumbar (lower back) segment of the vertebral column. It distributes sensory information from the body to the brain and CNS commands from the brain to the body, and also contains networks that control reflexes and central pattern generators. Graphic © Silver Place/Shutterstock.com.
The peripheral nervous system (PNS) consists of neurons and nerves outside of the brain and spinal cord and is comprised of the autonomic nervous system and somatic nervous system. Peripheral nervous system graphic adapted from © Elena Ladanovskaya/Shutterstock.com.
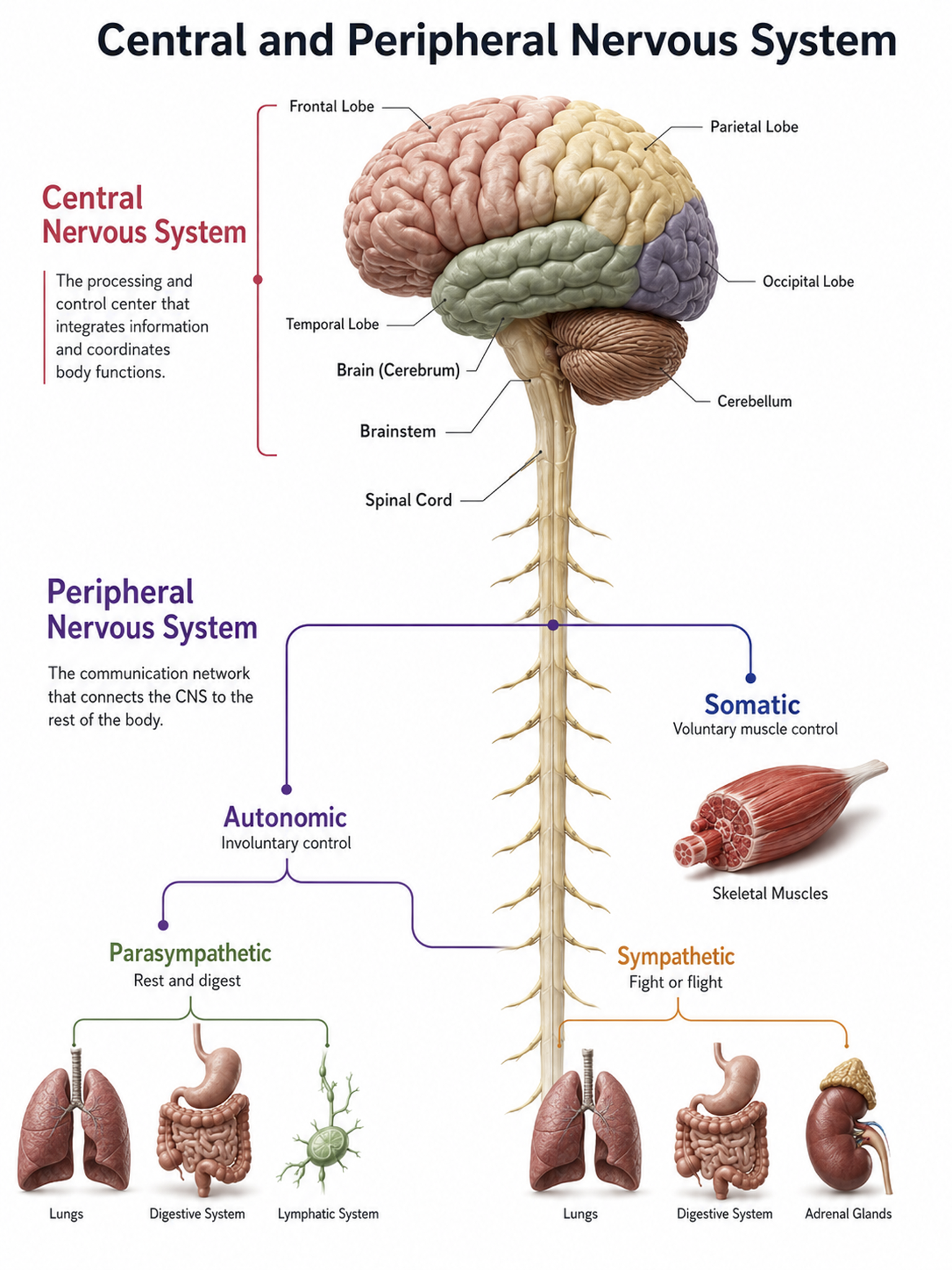
Nerves
Nerves are bundles of axons that lie outside of the central nervous system. Motor nerves distribute instructions from the CNS to the rest of the body, while sensory nerves transmit information from sensory receptors to the CNS.
There are three major systems of nerves: cranial nerves, spinal nerves, and the autonomic nervous system.
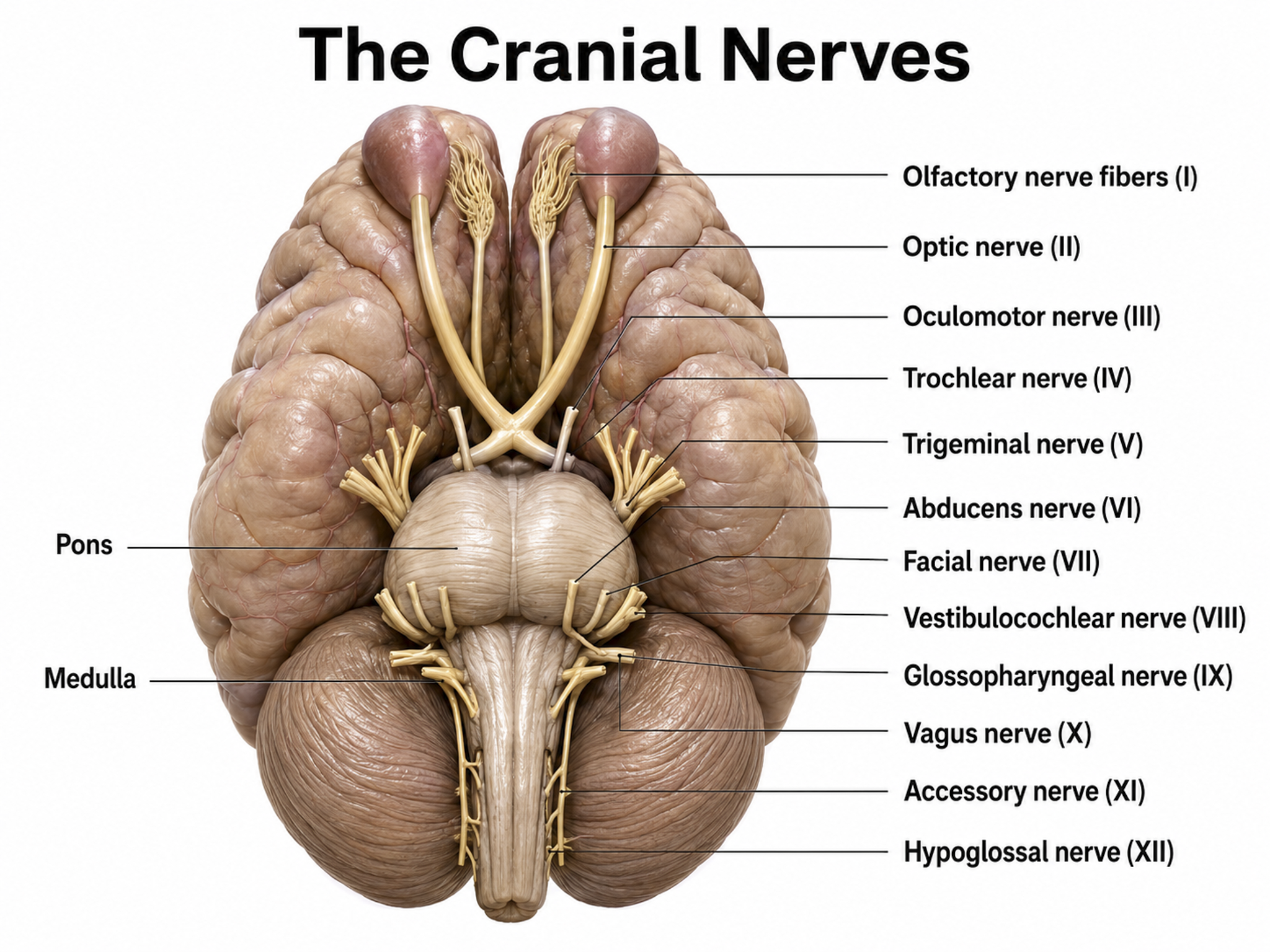
The 12 pairs of cranial nerves distribute sensory and motor information. Three are exclusively sensory pathways to the brain: olfactory (I), optic (II), and vestibulocochlear (VIII). Five are exclusively motor pathways from the brain: oculomotor (III), trochlear (IV), abducens (VI), spinal accessory (XI), and hypoglossal (XII). Four cranial nerves carry both sensory and motor information: trigeminal (V), facial (VII), glossopharyngeal (IX), and vagus (X). Graphic adapted from © Alila Medical Media/Shutterstock.com.
Thirty-one pairs of spinal nerves, each member serving one side of the body, leave the spinal cord through openings in the backbone. Spinal nerve graphic © SciePro/Shutterstock.com.
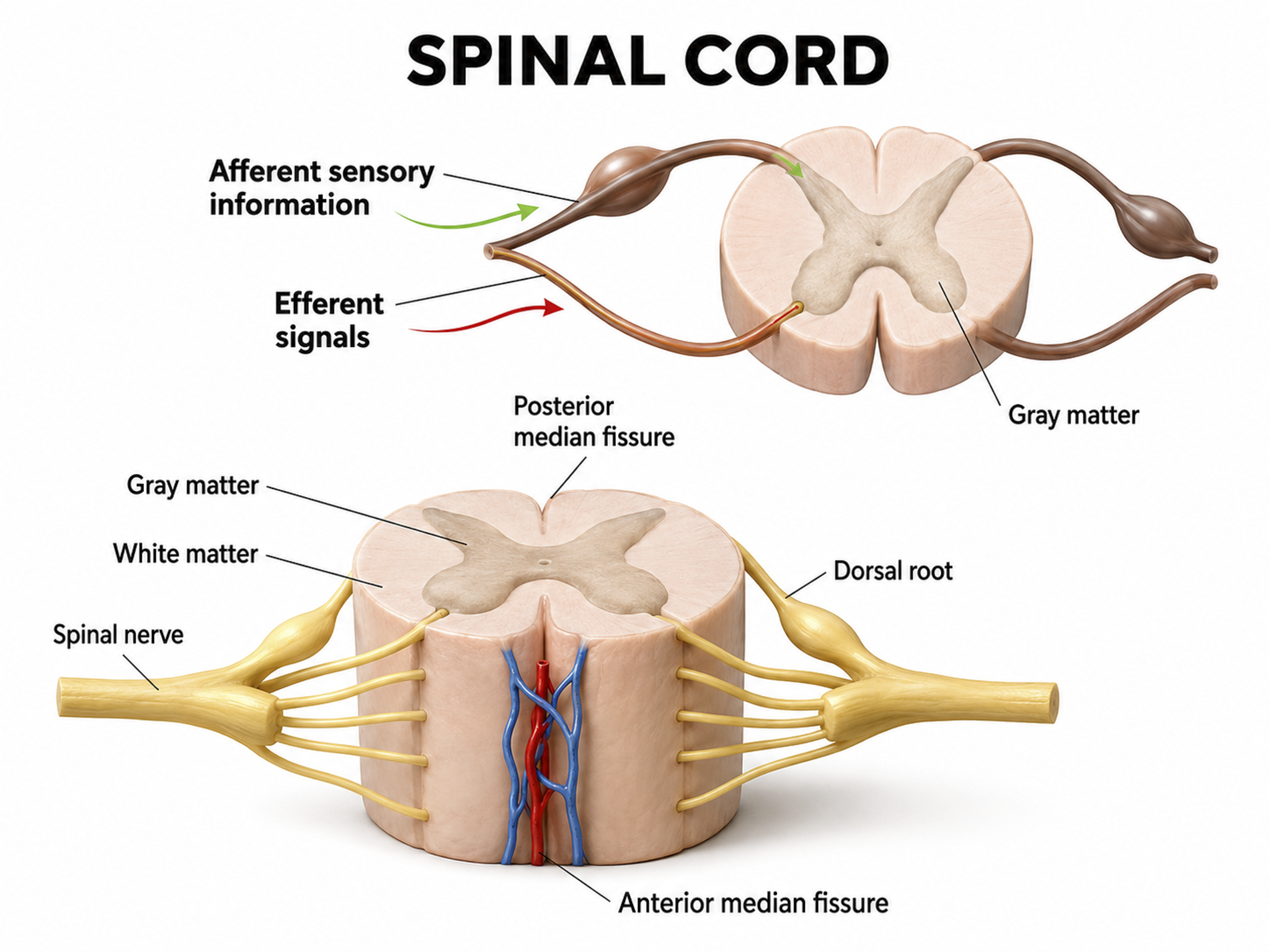
Each spinal nerve carries sensory projections from the body (dorsal root) and motor commands from the spinal cord to skeletal muscles (ventral root). Graphic adapted from © Designua/Shutterstock.com.
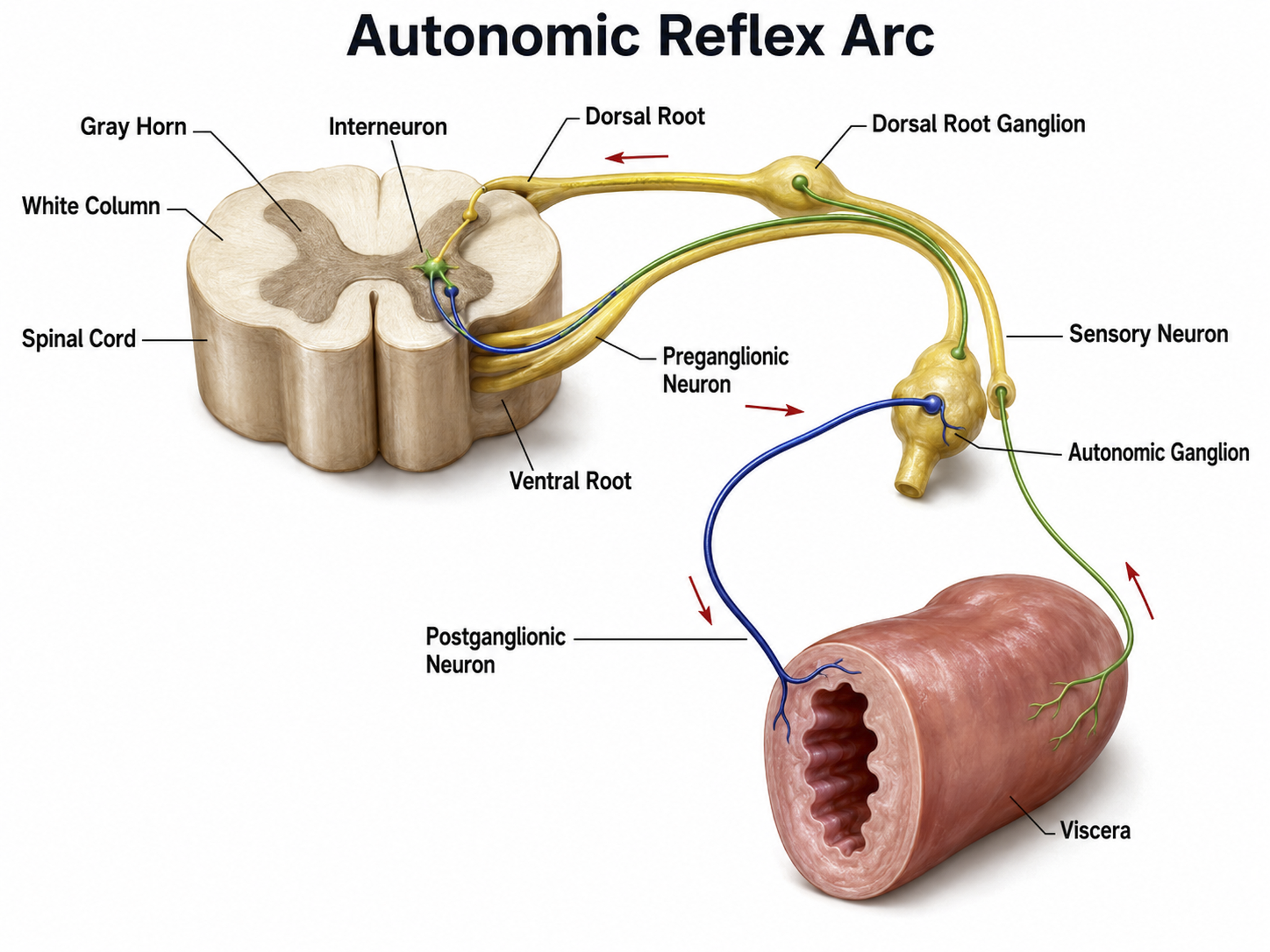
Autonomic Nervous System
The autonomic nervous system regulates cardiac and smooth muscle and glands, transmits sensory information to the CNS, and innervates muscle spindles. It is the brain's main system for monitoring and controlling major organs.
While we normally exercise little intentional, conscious autonomic control, self-regulation disciplines like yoga and extensive research in biofeedback have demonstrated that we can learn voluntary autonomic control. Clinicians can train clients to regulate heart rate variability, peripheral blood flow to the fingers, toes, and scalp, and finger sweat gland activity to treat disorders and achieve optimal performance.
The autonomic nervous system is divided into three main systems: sympathetic, parasympathetic, and enteric. Check out the YouTube video, The Autonomic Nervous System.
We activate the sympathetic nervous system when we encounter a threat that we can fight or flee, but we also activate this branch when we get up from the couch or exercise. The parasympathetic system can oppose or complement sympathetic activity. When we feel safe, it allows us to self-regulate (through meditation or neurofeedback skills), socially engage with others, and engage in executive functions like planning. When we feel endangered and cannot fight or flee, this system produces freezing, fainting, or dissociation responses.
The enteric system consists of over 100 million neurons that release over 30 neurotransmitters to control the gut under CNS supervision, helping to maintain fluid and nutrient balance.
Somatic Nervous System
The somatic nervous system is comprised of spinal nerves that innervate somatosensory receptors in the skin, joints, and skeletal muscles. While somatic motoneuron cell bodies lie in the CNS, most of their axons are in the PNS. The cell bodies of somatic sensory neurons are in the PNS dorsal root ganglia. Dorsal root graphic adapted from © stihii/Shutterstock.com.
Navigating the Brain
Orientations
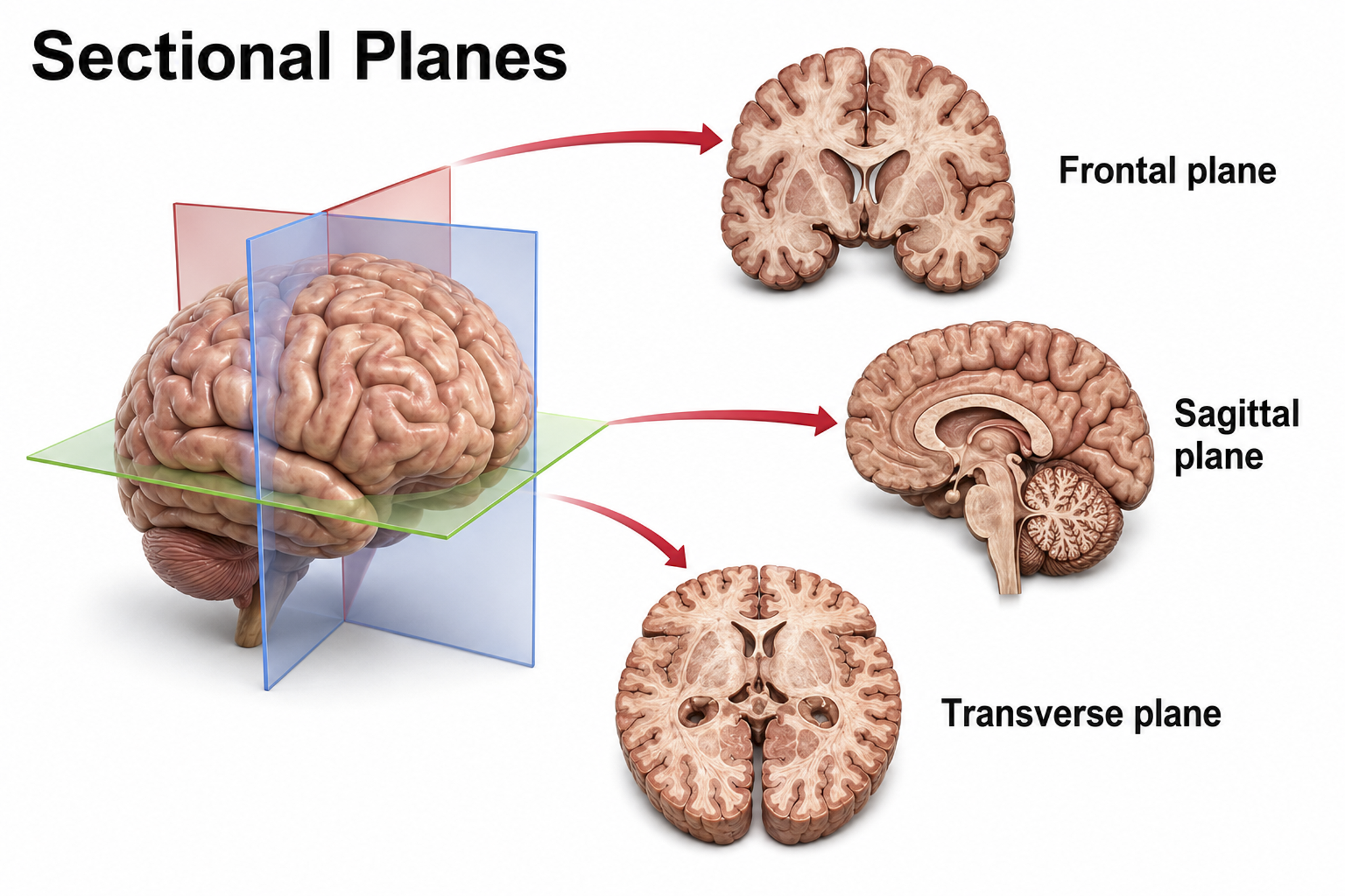
Three customary planes for viewing the body and brain are sagittal, coronal, and horizontal.
The sagittal plane divides the body into right and left halves. The coronal plane separates the body into front and back parts. The horizontal (transverse) plane divides the brain into upper and lower parts (Breedlove & Watson, 2023). These planes are used in neuroimaging and help clinicians interpret brain scans and localize areas of interest for neurofeedback training. Graphic adapted from Blausen.com staff "Blausen gallery 2014," Wikiversity Journal of Medicine.
Directional Terms
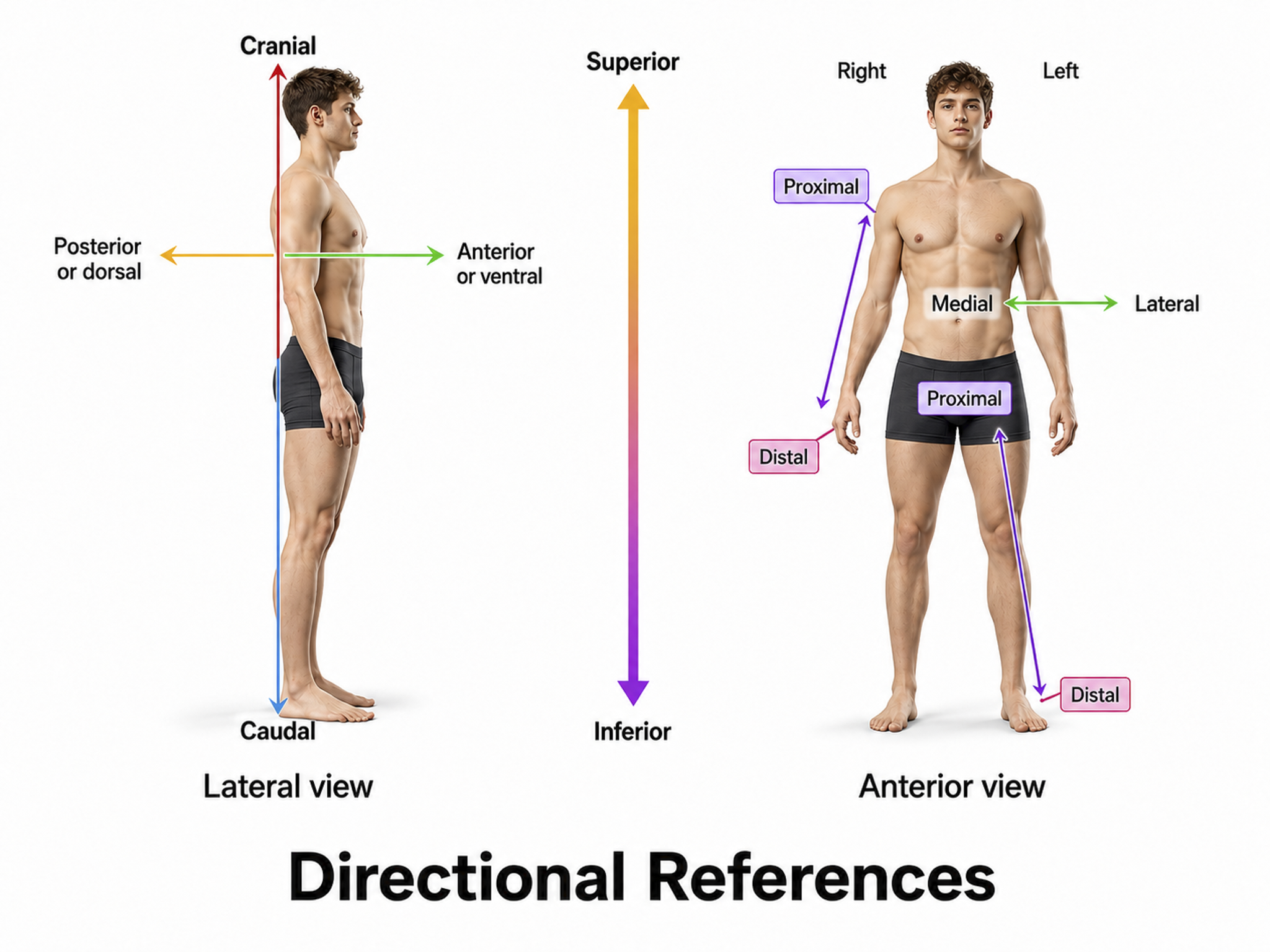
Important directional terms include medial (toward the middle) and lateral (toward the side), ipsilateral (same side) and contralateral (opposite side), superior (above) and inferior (below), anterior/rostral (toward the head) and caudal (posterior, toward or at the back), proximal (near the center) and distal (toward the periphery), and dorsal (toward or at the back) and ventral (toward the belly) (Breedlove & Watson, 2023). Graphic adapted from Blausen.com staff "Blausen gallery 2014," Wikiversity Journal of Medicine.
Cortical Features
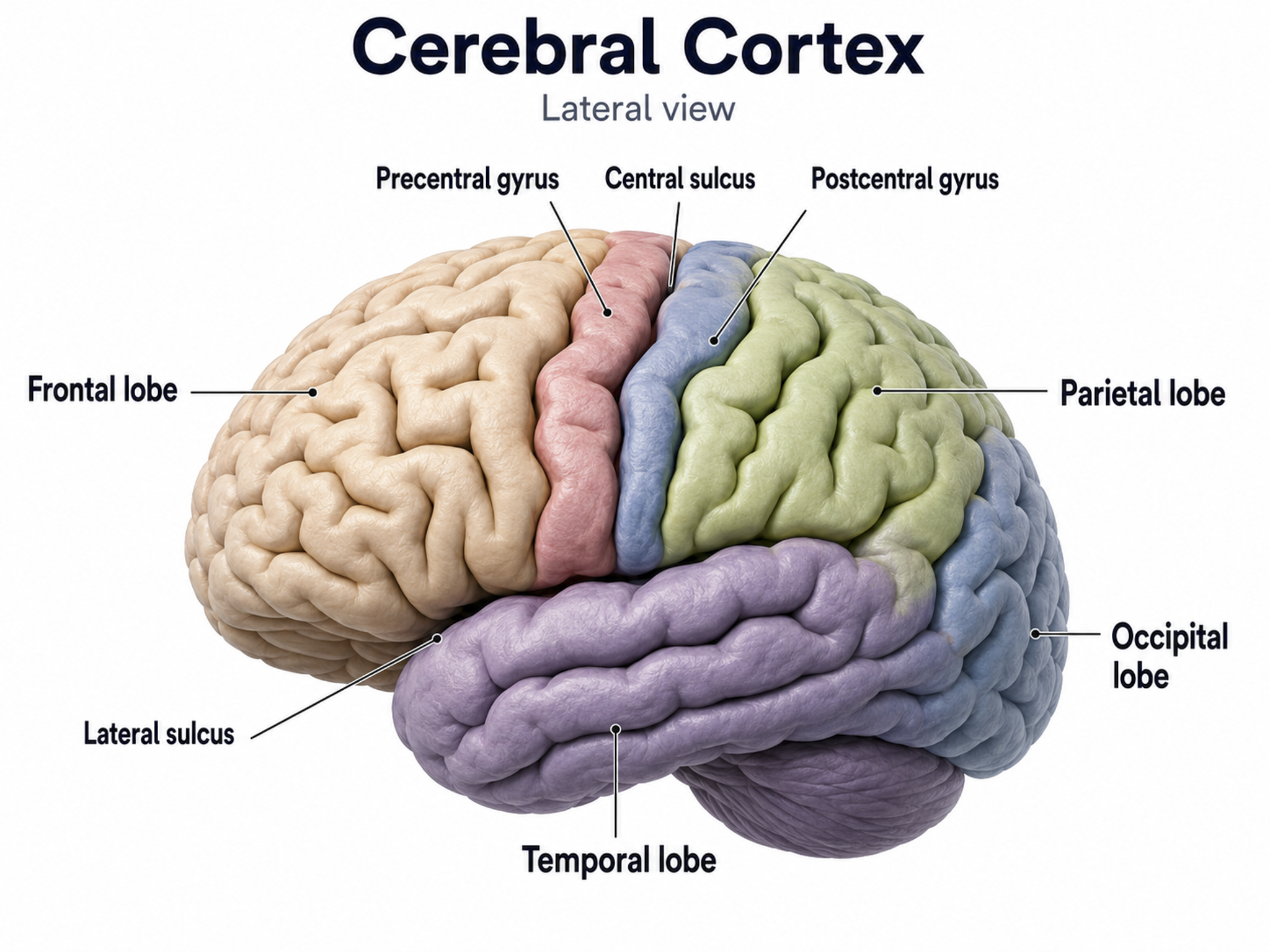
The adult human brain has a volume of roughly 1,100 to 1,300 cm3, varying with sex and body size, and requires convolutions to fit within the skull (Bear, Connors, & Paradiso, 2020). If the cortex were flattened into a sheet, its surface area would be about 2,000 to 2,500 cm2, roughly a quarter of a square meter or 2 to 2.7 square feet, with about two-thirds of the cortical surface lying within these folds (Breedlove & Watson, 2023). Anatomists distinguish three topographical features of the cerebral cortex: gyrus, sulcus, and fissure.
A gyrus is a ridged area of the brain. The precentral gyrus, anterior to the central sulcus, is the primary motor cortex (controlling muscles and movements). The postcentral gyrus, posterior to the central sulcus, is the primary somatosensory cortex (receiving somatosensory information).
A sulcus is a groove in the cortical surface. The central sulcus separates the primary motor cortex from the primary somatosensory cortex. A fissure is a deep groove; for example, the Sylvian fissure (also called the lateral fissure or lateral sulcus) forms the upper boundary of the temporal lobe (Breedlove & Watson, 2023). These features serve as reference landmarks for navigating brain anatomy.
The Unfixed Brain
This video was produced by Suzanne Stensaas, PhD, Department of Neurobiology and Anatomy, and the Spencer S. Eccles Health Sciences Library, University of Utah.
Dissecting Brains
This video is courtesy of the Wellcome Collection.
Clinical Application
Although we exercise little conscious control over the autonomic nervous system, disciplines like yoga and decades of biofeedback research show that voluntary regulation is possible. In practice, you can teach a client to raise heart rate variability, warm the fingers by increasing peripheral blood flow, or reduce finger sweat gland activity to manage a disorder or pursue optimal performance. Grounding these interventions in autonomic anatomy helps you explain to clients why the skills they practice produce measurable physiological change.
Key Takeaways
The central nervous system comprises the brain, spinal cord, and retina and contains roughly 86 billion neurons, while the peripheral nervous system divides into the autonomic and somatic systems. The body carries twelve pairs of cranial nerves and thirty-one pairs of spinal nerves that route sensory and motor information. The autonomic nervous system has sympathetic, parasympathetic, and enteric divisions, and biofeedback demonstrates that its activity can be voluntarily influenced. Anatomical planes, sagittal, coronal, and horizontal, together with directional terms, orient neuroimaging and electrode placement. The cortex is heavily folded, with the precentral gyrus serving as primary motor cortex and the postcentral gyrus as primary somatosensory cortex, separated by the central sulcus.
Check Your Understanding
- Name the three structures that make up the central nervous system.
- How many cranial nerve pairs are exclusively sensory, exclusively motor, and both, and give one example of each?
- Which anatomical plane divides the body into right and left halves, and which divides it into front and back?
- Which gyrus contains the primary motor cortex, which contains the primary somatosensory cortex, and what separates them?
Lateral and Medial Brain Landmarks
Subdivisions of the Brain
The brain is divided into three major subdivisions: forebrain, midbrain, and hindbrain. Brain landmark graphic adapted from © snapgalleria/Shutterstock.com.
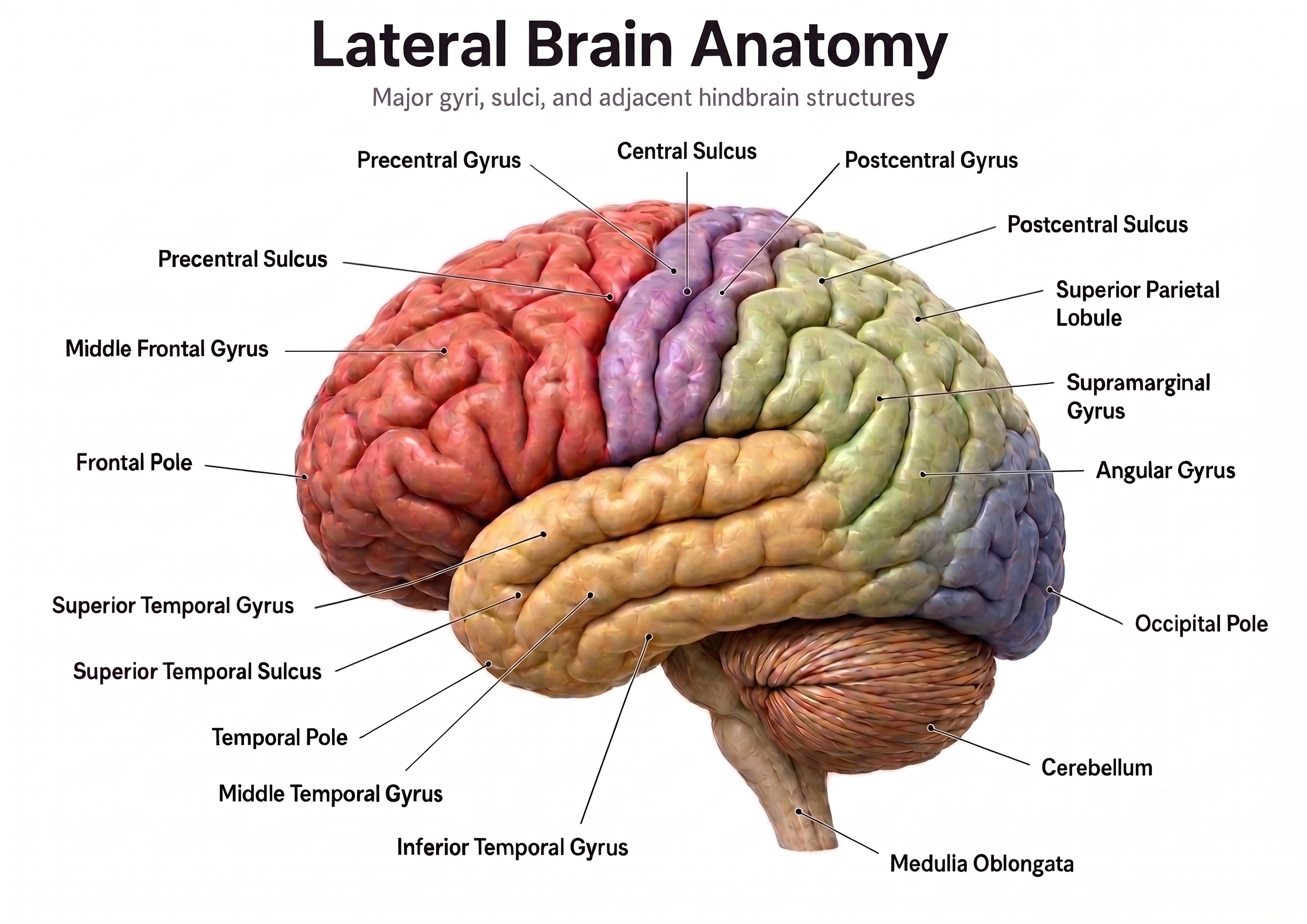
Medial brain section graphic adapted from © NatthapongSachan/Shutterstock.com.
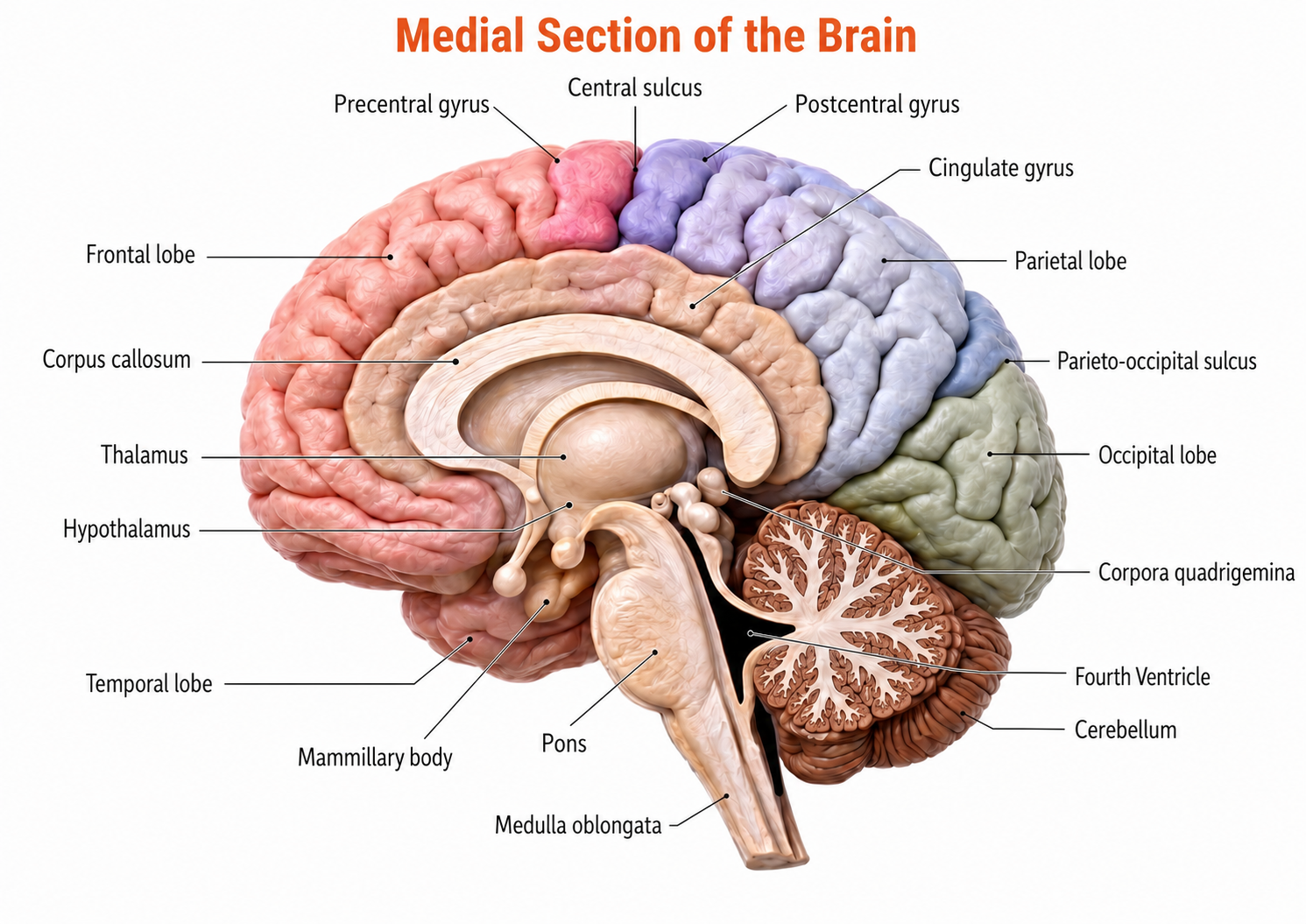
The forebrain consists of the telencephalon (cerebral hemispheres) and the diencephalon. The telencephalon encompasses the cerebral cortex and the deeper structures of the basal ganglia and limbic system. Limbic system graphic © SciePro/Shutterstock.com.
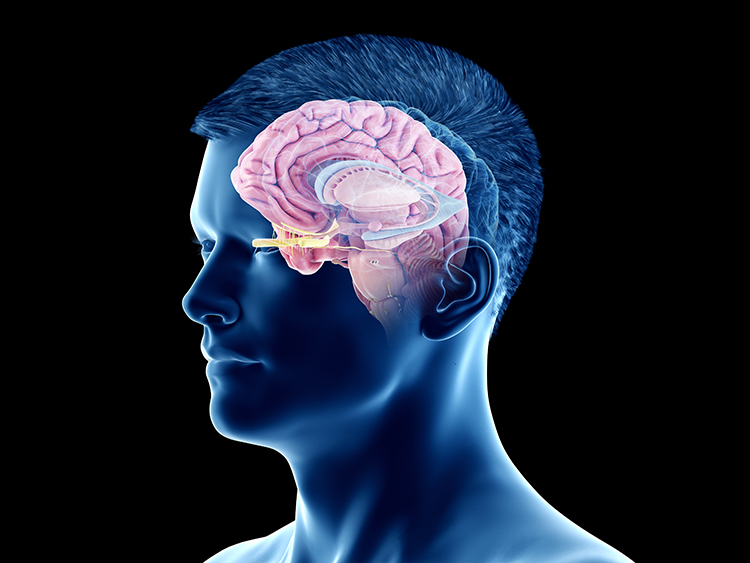
The diencephalon in the posterior forebrain contains the thalamus and hypothalamus. Thalamus graphic © SciePro/Shutterstock.com.
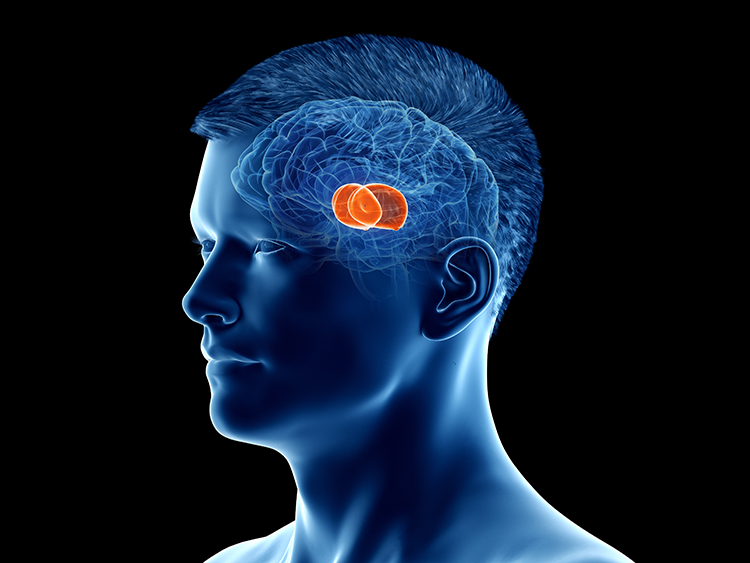
The midbrain consists of the mesencephalon, which includes the inferior colliculi, superior colliculi, and substantia nigra. The degeneration of the substantia nigra is a key step in developing Parkinson's disease. Substantia nigra graphic © Kateryna Kon/Shutterstock.com.
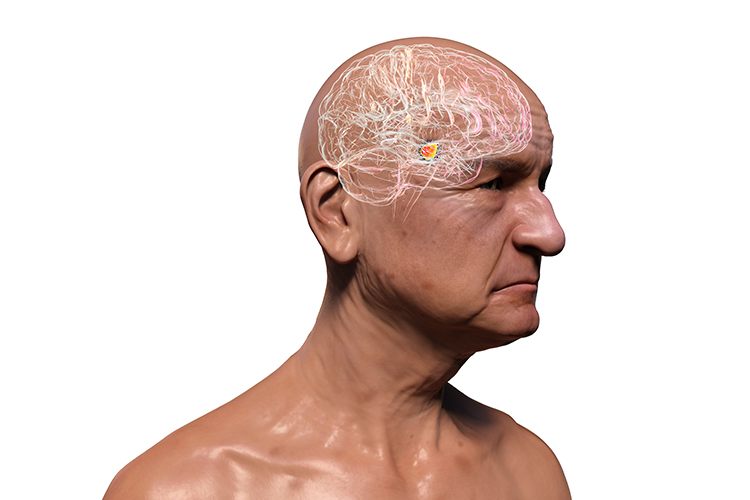
The hindbrain contains the metencephalon and myelencephalon. The metencephalon is comprised of the cerebellum and pons. The cerebellum plays a role in higher-level functions like emotional and cognitive regulation, influencing the speed, capacity, consistency, and appropriateness of cognitive and emotional processes. Damage to the cerebellum can impair cognitive and affective performance as well as movement.
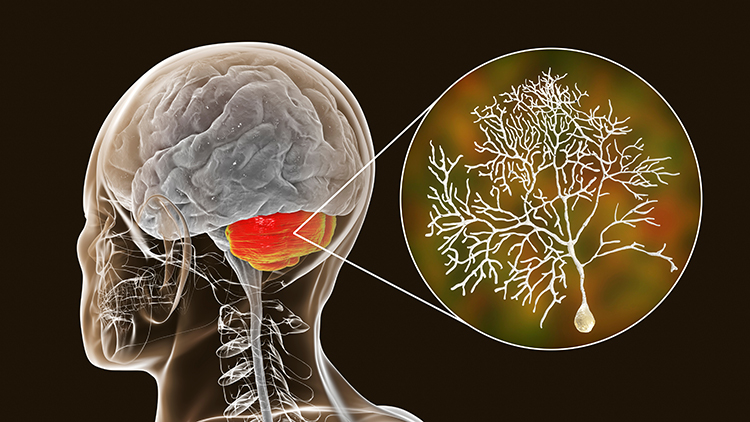
The cerebellum provides coordination and fine-tuning of balance and movement, addressing the rate, rhythm, and force of movement as well as analogous qualities in thinking and emotional expression. Cerebellum graphic with highlighted Purkinje neuron © Kateryna Kon/Shutterstock.com.
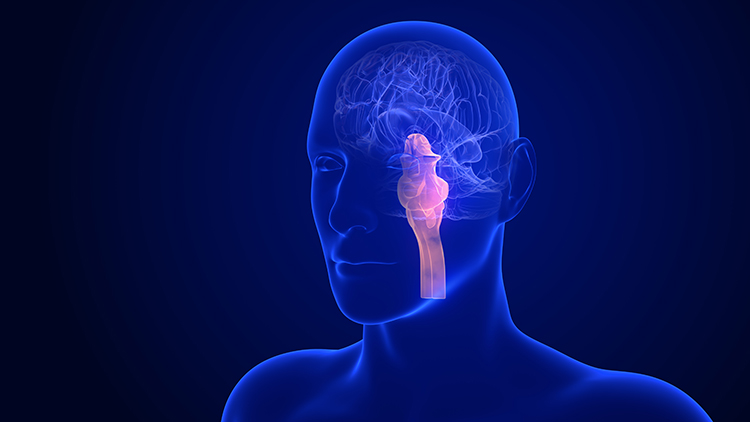
The myelencephalon consists of the medulla, which plays a critical role in the speeding and slowing of the heart across each breathing cycle, a phenomenon called respiratory sinus arrhythmia (RSA). This process is a key target in heart rate variability (HRV) biofeedback training. Alcohol, opioids, and sedative-hypnotics can fatally depress brainstem respiratory centers, slowing and halting breathing. Medulla graphic © mkfilm/Shutterstock.com.
The Skull
The human skull is a complex anatomical structure that plays a significant role in protecting the brain and influencing EEG signal measurement. Skull anatomy graphic adapted from © Magic mine/Shutterstock.com.
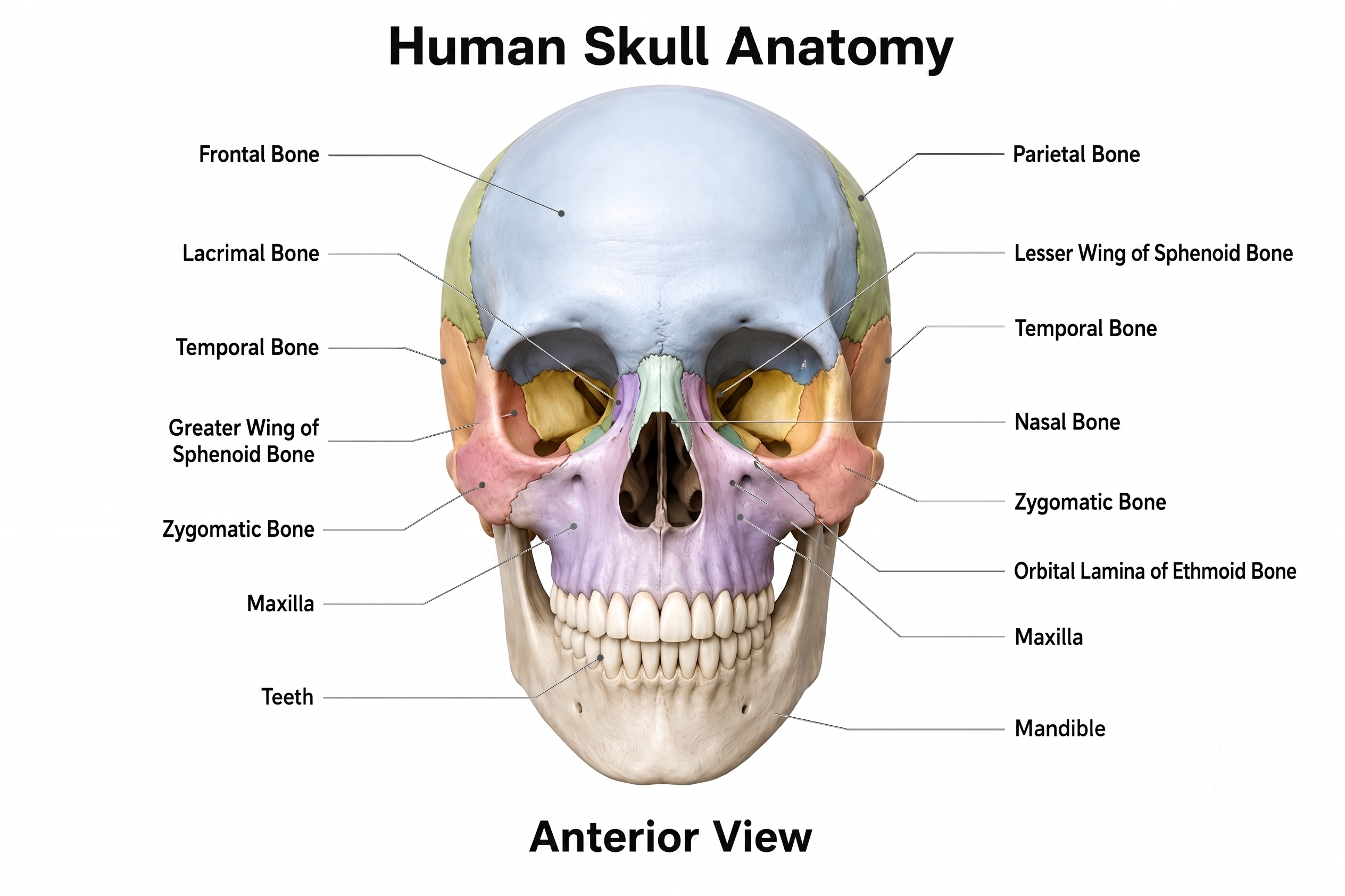
Comprising 22 bones, the skull is divided into the cranium, which houses the brain, and the facial bones. The cranium is composed of several major bones, including the frontal, parietal, temporal, and occipital bones, fused together by sutures. This bony framework provides a rigid protective case for the brain while also anchoring the meninges and other protective layers (Niedermeyer & Lopes da Silva, 2004).
The skull's primary function is to protect the brain from physical damage. Its bones have an average thickness of about 6.5 mm in adults, which significantly impacts EEG signal transmission. The skull acts as a low-pass filter, attenuating high-frequency components of the brain's electrical activity while allowing lower-frequency components to pass through. The skull can reduce signal amplitude by approximately 85% (He & Li, 2010), primarily due to its low conductivity compared to brain and scalp tissues. This filtering effect is a crucial consideration in EEG signal analysis and interpretation (Sanei & Chambers, 2013).
Key Concept
The skull behaves as a low-pass filter, attenuating the high-frequency components of the brain's electrical activity while allowing lower frequencies to pass. Its low conductivity can reduce EEG signal amplitude by roughly 85 percent, and its mix of dense and porous bone produces anisotropic attenuation that varies with the direction of current flow. These properties mean that much of the neural signal is dissipated and distorted before it reaches a scalp electrode. Accounting for skull filtering is essential when interpreting amplitudes and when using source localization or connectivity techniques.
For example, if a skull defect leaves an opening, then fast-frequency EEG will be seen over the defect (i.e., breach rhythm).
The skull's high impedance means that much of the electrical activity generated by neurons is dissipated before reaching scalp electrodes, making it more challenging to detect subtle neural oscillations. Additionally, the varying thickness and composition of skull bones can introduce spatial distortions in EEG signals, complicating the localization of neural activity sources (Lopes da Silva, 2010).
The skull's heterogeneous structure, including both dense cortical bone (dense bone that forms a cortex or “bark” around the bone) and less dense cancellous bone (porous bone in the marrow-filled medullary cavity of bones), causes anisotropic attenuation, meaning that the degree of signal reduction varies depending on the direction of electrical currents (i.e., the skull conducts current more readily along some directions than others, so the degree of attenuation depends on the orientation of the current path and not on distance alone). This anisotropy must be accounted for in advanced EEG analysis techniques such as source localization and brain connectivity studies (Nunez & Srinivasan, 2006).
In addition to the bones, the skull's sutures and foramina (openings) influence EEG signal propagation. Sutures can act as additional barriers or channels for electrical signals, while foramina may create localized points of less attenuation. Individual variability in skull anatomy also significantly influences EEG signals: children's skulls are generally thinner and less dense than those of adults, potentially resulting in less signal attenuation (Schoffelen & Gross, 2009). The scalp and meninges add further layers of impedance that electrical signals must traverse, each with distinct electrical properties. Understanding the combined effect of these tissues is essential for accurate EEG interpretation and for the development of advanced EEG techniques (Niedermeyer & Lopes da Silva, 2004).
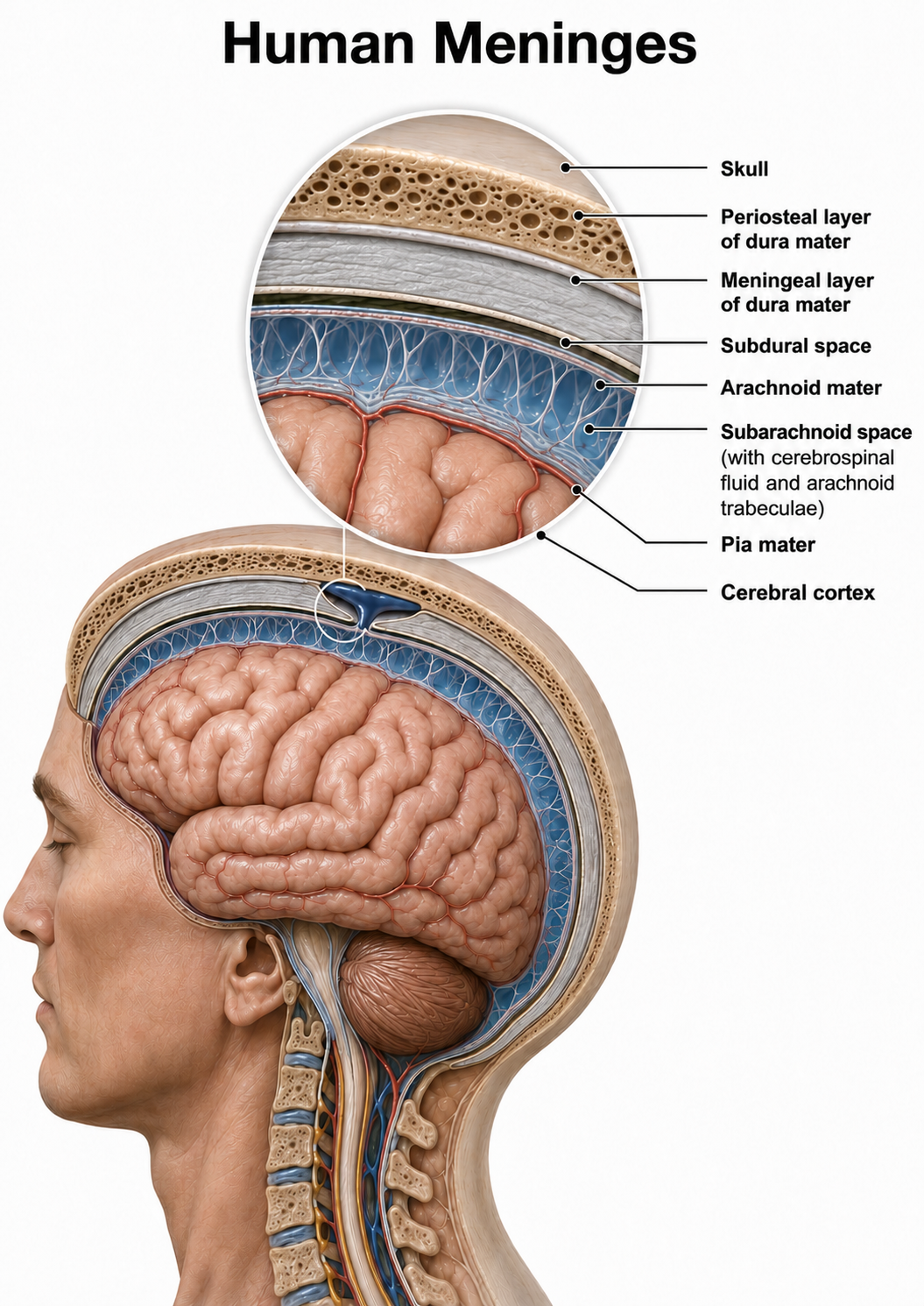
Meninges
Three meninges protect the brain and spinal cord, which are housed within the skull and vertebrae. The choroid plexus of the ventricles produces cerebrospinal fluid, which then circulates through the ventricles and the subarachnoid space of the meninges. These membranes include the dura mater, pia mater, and middle arachnoid (Breedlove & Watson, 2023). Graphic adapted from © Alilia Medical Media/Shutterstock.com.
Cerebral Ventricles
The cerebral ventricles are a network of fluid-filled chambers that protect the brain from trauma due to abrupt head movements and facilitate the exchange of nutrients and wastes between blood vessels and the brain. These cavities, found within all four lobes of each hemisphere, include the lateral, third, and fourth ventricles.
The brain contains four interconnected ventricles. A pair of lateral ventricles occupies the two cerebral hemispheres, the third ventricle sits in the midline between the left and right halves of the thalamus and hypothalamus, and the fourth ventricle lies between the pons and upper medulla in front and the cerebellum behind. Narrow channels connect these cavities: the paired interventricular foramina (foramen is the singular, meaning opening) join each lateral ventricle to the third ventricle, and the cerebral aqueduct joins the third ventricle to the fourth (Breedlove & Watson, 2023). Graphic adapted from © joshya/Shutterstock.com.
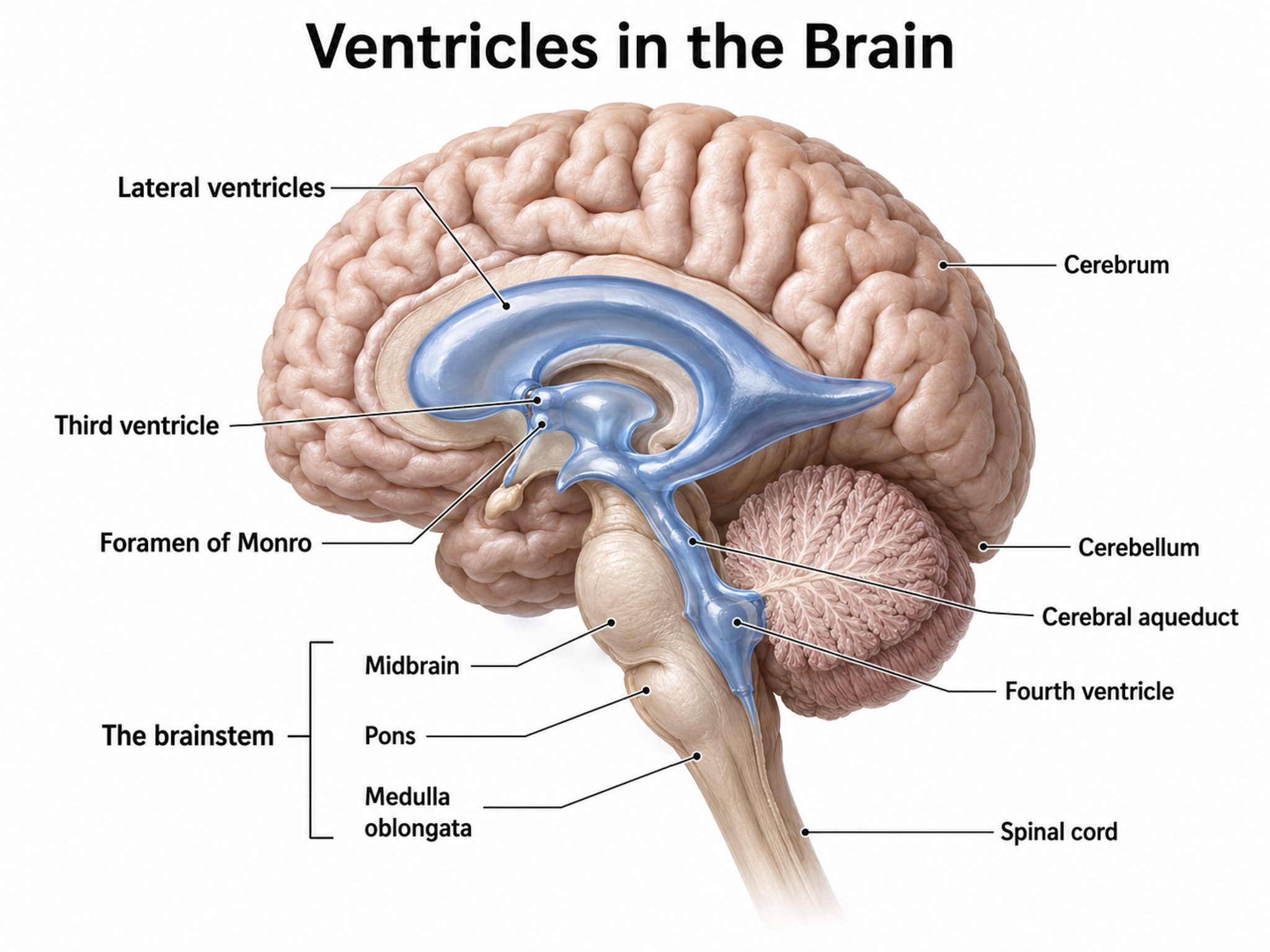
Glymphatic System
The glymphatic system is a recently discovered lymphatic-like system in the brain. It provides a flow of CSF through the brain's interior that helps clear cellular debris, proteins, and other wastes. Glymphatic system graphic © Claus Lunau/Science Photo Library.
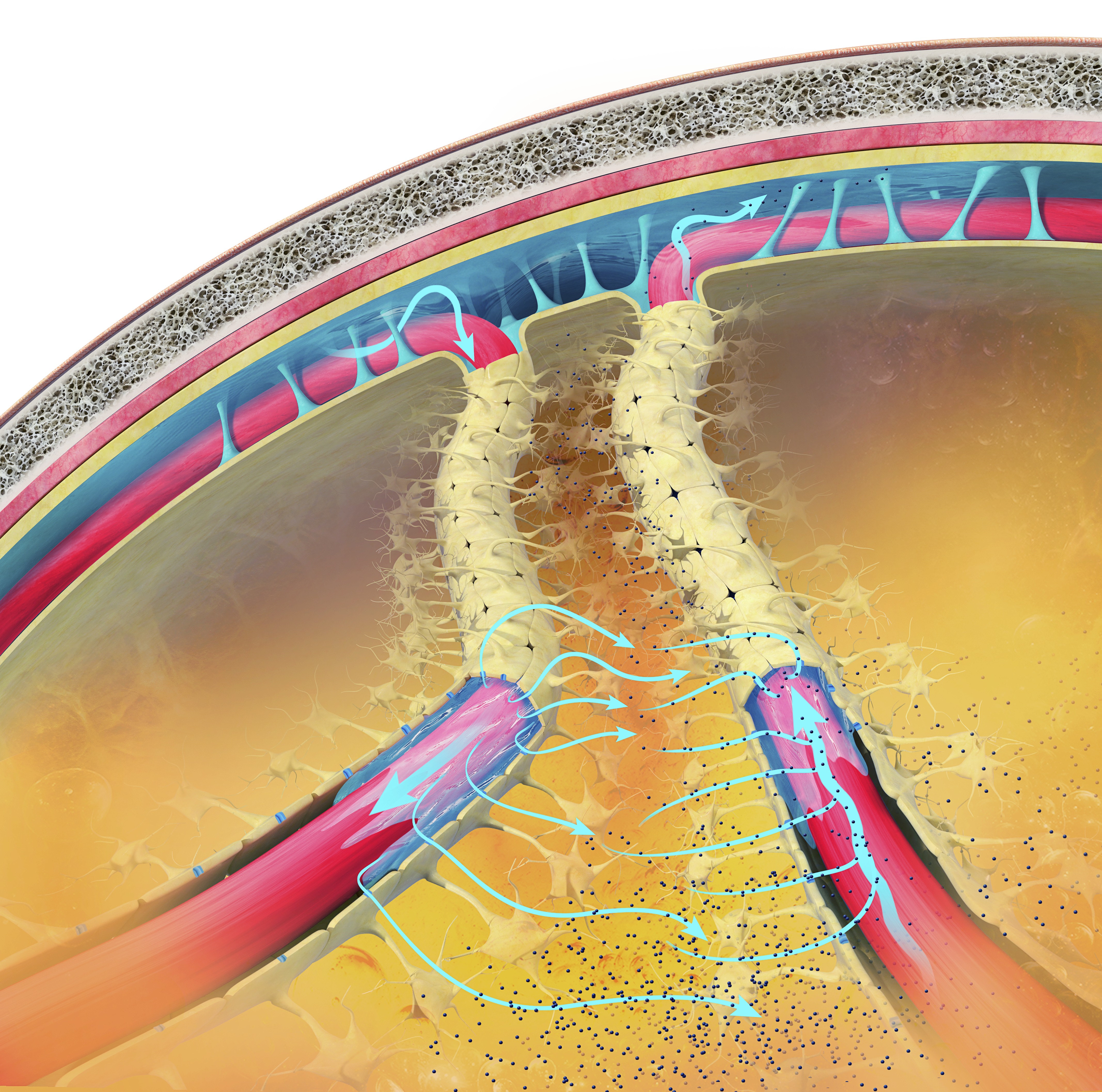
Also known as the glymphatic clearance pathway or the paravascular system, this system clears waste and fluid from the vertebrate central nervous system (CNS). Interstitial fluid is removed via the cerebrospinal fluid (CSF). It is similar to the lymphatic system, but functions to remove waste products from the brain and spinal cord. This view shows the subarachnoid space (across top) between the brain and its membranes. The blue arrows show the movement of interstitial fluid and solutes.
By removing harmful substances such as the amyloid-beta and tau proteins implicated in Alzheimer's disease and the alpha-synuclein implicated in Parkinson's disease, the glymphatic flow may protect us from various neurological disorders (Breedlove & Watson, 2023). The glymphatic system removes most of its waste during stage-3 sleep (slow-wave sleep), which underscores the clinical importance of healthy sleep, a factor biofeedback practitioners should assess in every client.
The Brain's Vascular System
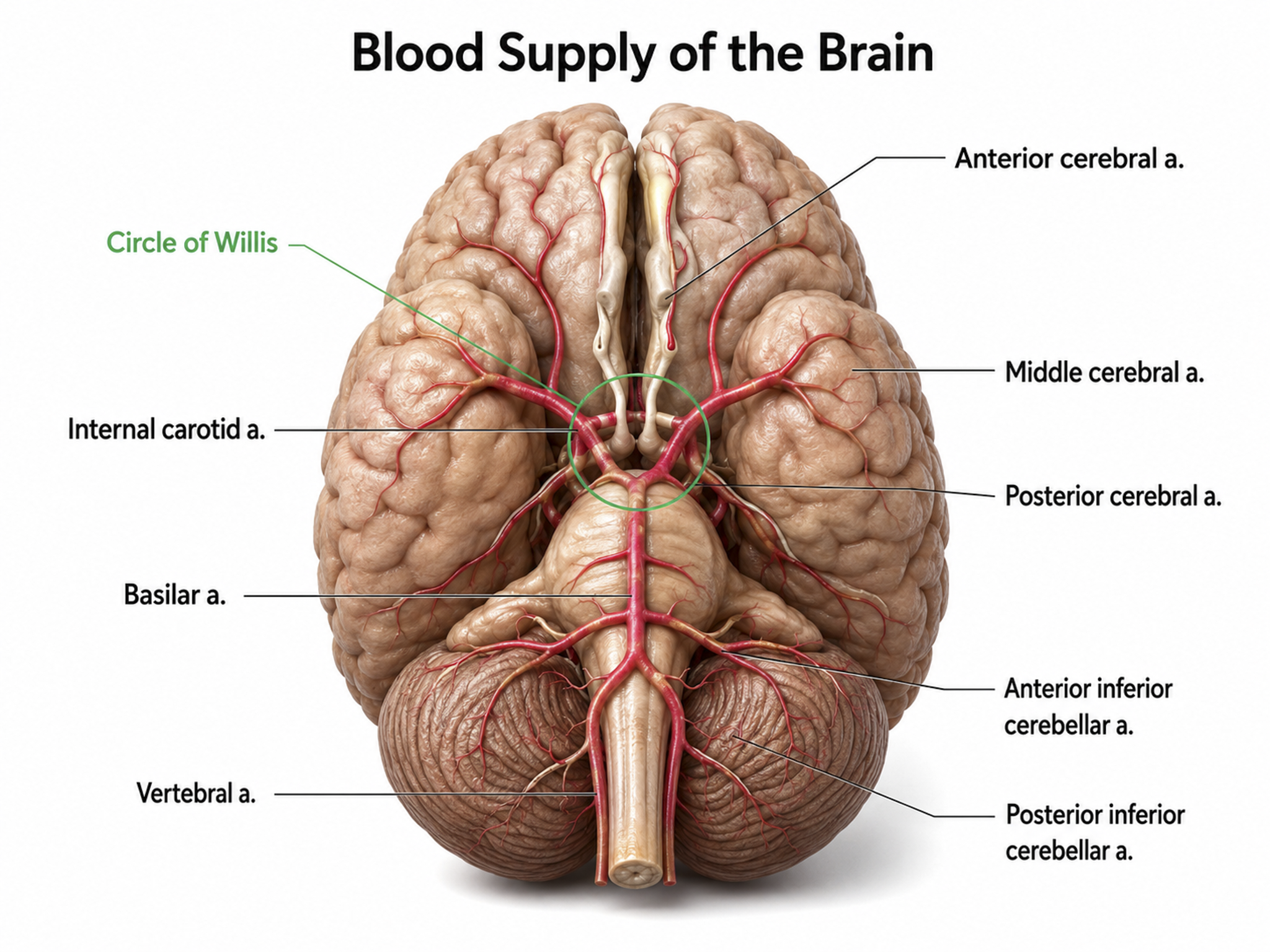
The resting brain consumes over 20% of the body's energy. The internal carotid artery's anterior and middle cerebral arterial branches deliver blood to about two-thirds of the cerebral hemispheres, while the paired vertebral arteries merge to form the basilar artery, whose left and right posterior cerebral arterial branches supply blood to the posterior cerebral hemispheres, cerebellum, and brainstem (Breedlove & Watson, 2023).
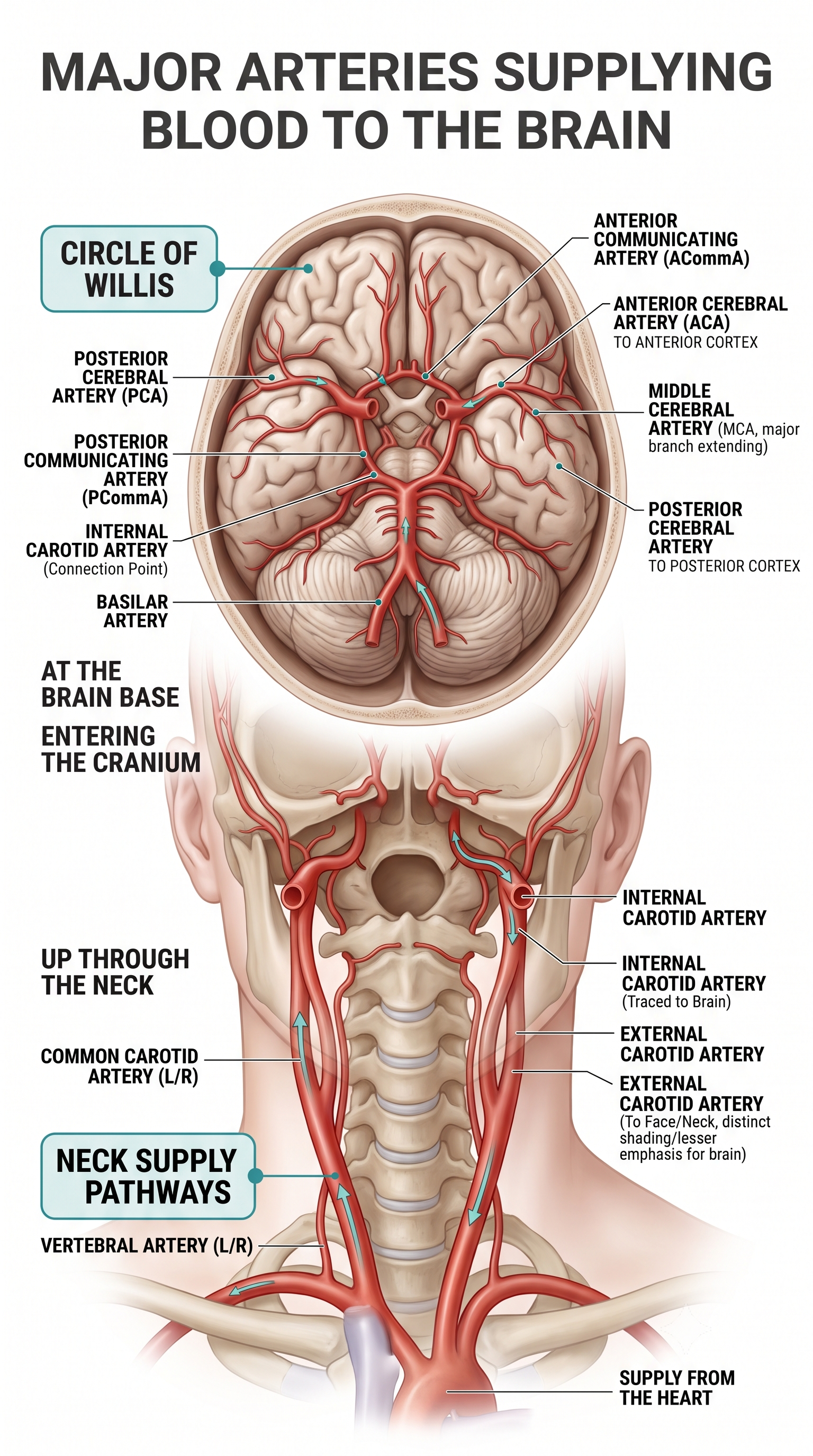
The effects of a stroke due to blood vessel blockage or rupture may be limited because paired arteries supply each brain hemisphere. The circle of Willis, a vascular network at the base of the brain comprised of the carotid and basilar arteries, may provide an alternate route for delivering blood when a major artery is compromised by disease or traumatic injury. The veins and sinuses drain deoxygenated blood, completing the circulatory loop to the heart via the jugular vein. Graphic adapted from © Alilia Medical Media/Shutterstock.com.
The figure below shows areas of the cortex that may be affected by strokes affecting the three major cerebral arteries.
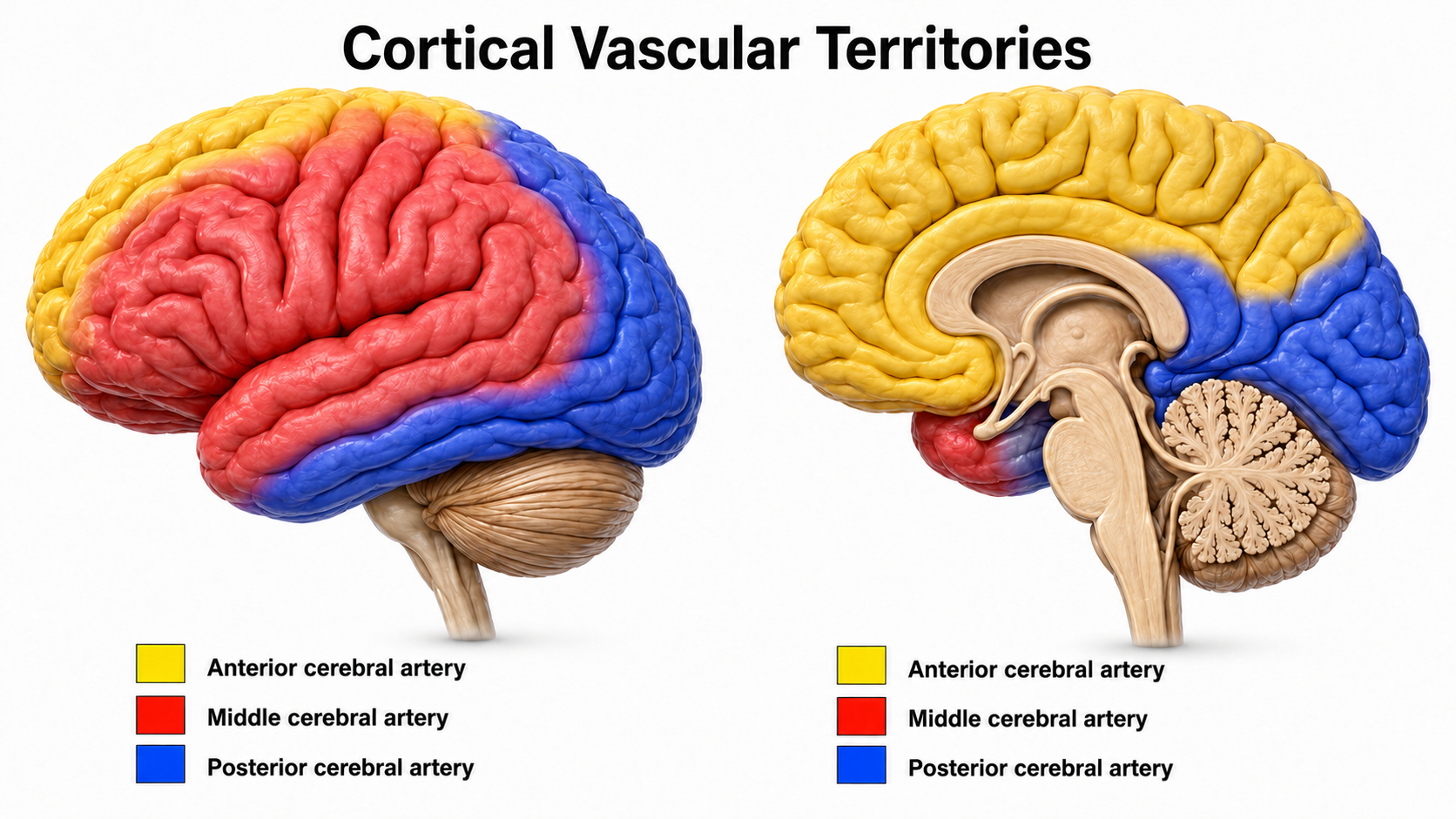
Graphic adapted from Cruces et al. (2022).
After perfusing the brain, blood depleted of oxygen, glucose, and other nutrients drains into a network of cerebral veins that differs strikingly from the venous anatomy found elsewhere in the body. Unlike most veins, cerebral veins are thin-walled, lack valves, and do not travel alongside their companion arteries, so drainage depends on pressure gradients and posture rather than on one-way valves. Superficial cortical veins collect blood from the outer surface of the cerebrum and empty into the dural venous sinuses, most notably the superior sagittal sinus, while deep veins gather blood from the basal ganglia, thalamus, and deep white matter and channel it through the great cerebral vein of Galen into the straight sinus. These sinuses converge at the confluence of the sinuses, then course through the transverse and sigmoid sinuses before exiting the skull as the internal jugular veins, which carry the blood toward the superior vena cava and the right atrium.
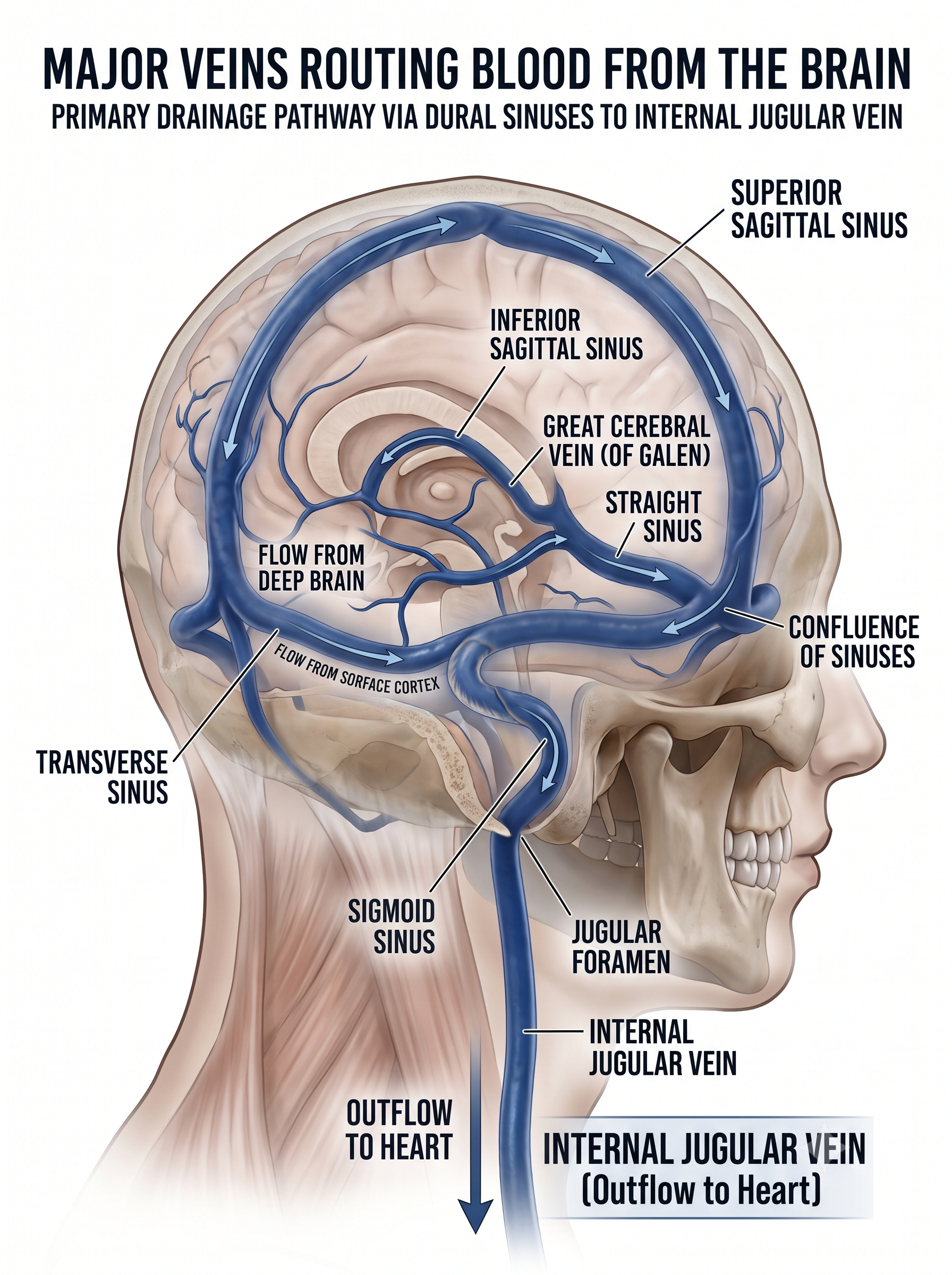
The same dural sinuses also house meningeal lymphatic vessels, a discovery that overturned the long-held belief that the central nervous system lacked lymphatic drainage and now links cerebral venous anatomy to waste clearance and immune surveillance in the brain (Aspelund et al., 2015; Louveau et al., 2015).
Blood-Brain Barrier
The blood-brain barrier (BBB) is a highly selective, semipermeable interface formed by the endothelial cells that line the brain's microvasculature, where continuous tight junctions seal the spaces between adjacent cells and force most substances to move through the cells rather than between them. This barrier is not the work of endothelial cells alone but of a coordinated neurovascular unit that also includes pericytes embedded in the capillary wall, the endfeet of astrocytes that ensheath the vessels, microglia, and the surrounding basement membrane.
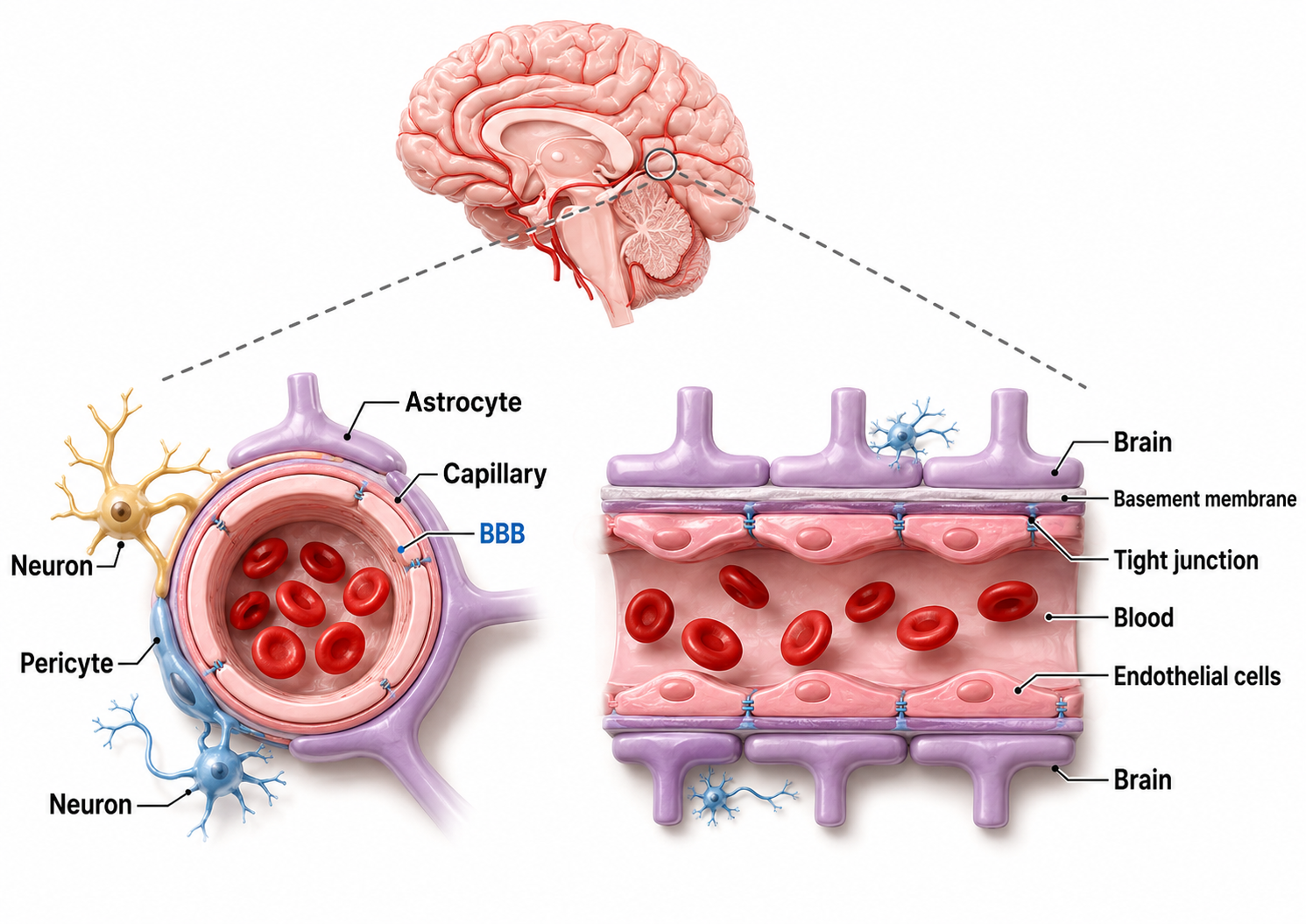
Image adapted from Ding et al. (2025).
Together these components induce and maintain the barrier's low permeability, minimal pinocytosis, and dense expression of transport proteins that set brain capillaries apart from those elsewhere in the body (Abbott et al., 2010). The result is a living border that behaves less like a passive wall and more like a tightly regulated gatekeeper for the central nervous system (Daneman & Prat, 2015).
Functionally, the BBB shields neural tissue from circulating pathogens, toxins, and swings in blood composition while precisely controlling the traffic of ions, nutrients, and signaling molecules that keep the neuronal microenvironment stable. Small lipid-soluble gases such as oxygen and carbon dioxide diffuse across freely, whereas glucose, amino acids, and other essential polar molecules depend on specific carrier and receptor-mediated systems, and efflux pumps actively return many foreign compounds to the blood (Abbott et al., 2010).
This same protective selectivity is a double-edged property, because it excludes the vast majority of small-molecule drugs and nearly all large-molecule therapeutics, making the BBB one of the central obstacles in treating neurological and psychiatric disorders. When the barrier breaks down, as it does in conditions ranging from stroke and multiple sclerosis to Alzheimer's disease, the loss of this regulation contributes directly to disease progression, which is why BBB integrity remains a priority in both neuroscience and drug development (Daneman & Prat, 2015).
Interim Summary
The nervous system's architecture (from its major divisions to the protective skull, meninges, and vascular supply) directly affects how we record and interpret brain signals. Skull thickness, meningeal layers, and vascular dynamics all influence the EEG data you collect in clinical sessions.
Key Takeaways
The brain divides into the forebrain, which contains the telencephalon and diencephalon, the midbrain, and the hindbrain, which houses the cerebellum, pons, and medulla. The skull acts as a low-pass filter that can reduce EEG amplitude by roughly 85 percent and introduces anisotropic distortion that varies with current direction. Three meninges and the cerebrospinal fluid produced by the choroid plexus cushion the brain, while the glymphatic system clears wastes such as amyloid-beta, tau, and alpha-synuclein mainly during slow-wave sleep. Paired cerebral arteries and the circle of Willis supply the brain and can limit stroke damage by providing alternate routes. The blood-brain barrier, now understood as a coordinated neurovascular unit, tightly regulates what reaches neural tissue and excludes most therapeutic drugs.
Check Your Understanding
- List the three major subdivisions of the brain and name one structure in each.
- Why is the skull described as a low-pass filter, and approximately how much does it reduce EEG signal amplitude?
- What is a breach rhythm, and what skull condition produces it?
- When is the glymphatic system most active, and why does this matter clinically?
- What is the neurovascular unit, and why does the blood-brain barrier complicate drug treatment of neurological disorders?
Major Functions of Cortical Lobes and Major Subcortical Structures and Brodmann Areas
This section surveys the functional roles of the brain's major cortical lobes, the insular cortex, the mirror neuron system, key subcortical structures, and the fiber tracts that connect them. For biofeedback practitioners, understanding these structure-function relationships is fundamental to interpreting EEG patterns and selecting training protocols.
Cortical Lobes
The cortical lobes are named for the overlying bones of the skull (Breedlove & Watson, 2023). Graphic © Sebastian Kaulitzki/Shutterstock.com.
The cortex is required for executive functions like attention, planning, and problem-solving.
Without a cerebral cortex, a person would be blind, deaf, dumb, and unable to initiate voluntary movement (Bear, Connors, & Paradiso, 2020, p. 205).
The five major cortical regions include the frontal, parietal, temporal, and occipital lobes, and the insula (not shown). Cortical lobes graphic adapted from © Madrock24/Shutterstock.com.
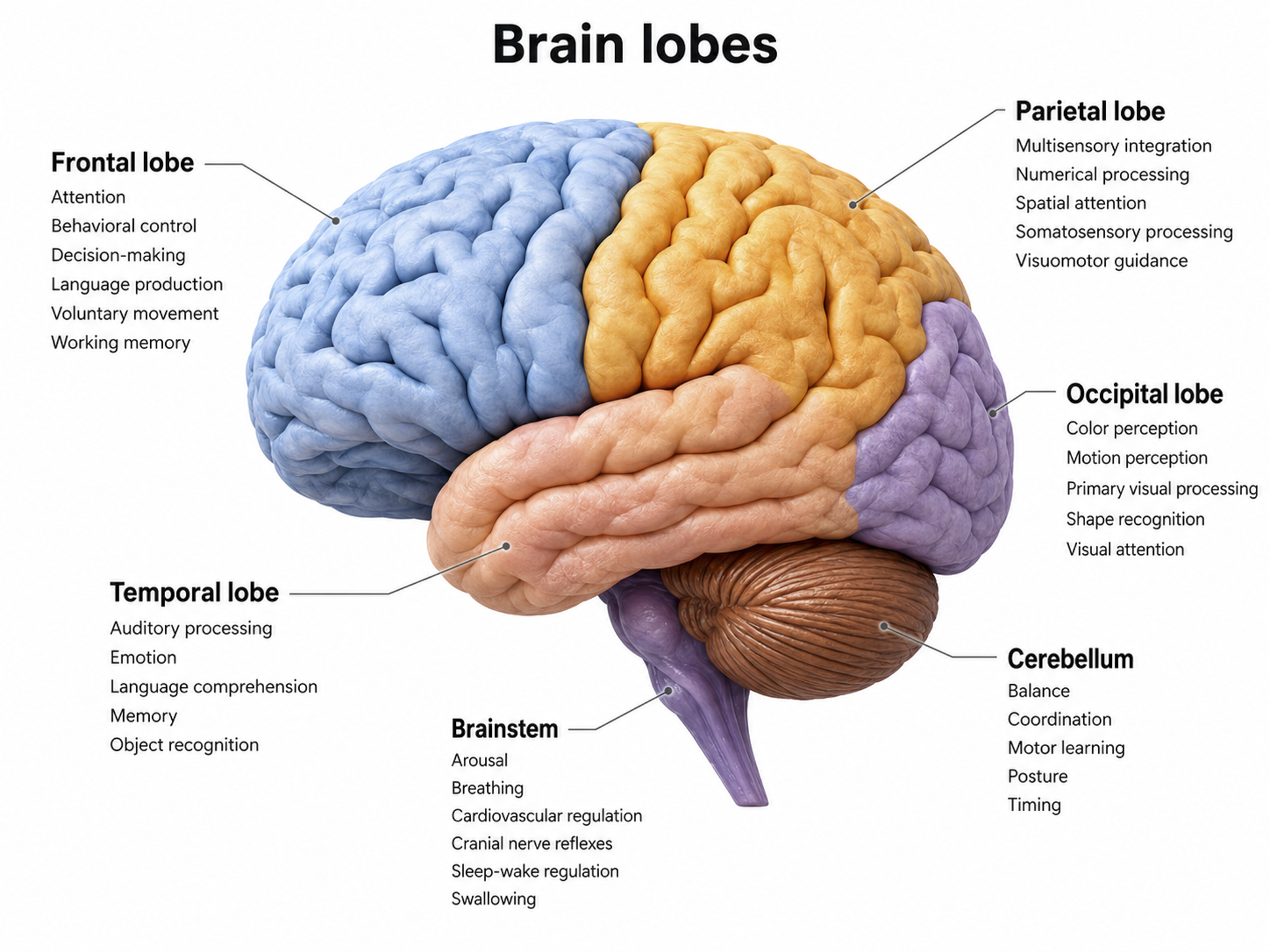
Frontal Lobes
The frontal lobes (Fp1, Fp2, F7, F3, Fz, F8, F4) consist of the cortex anterior to the central sulcus and include the primary motor cortex, motor association cortex, Broca's area, and prefrontal cortex. These structures collectively support everything from fine motor control to the highest-level executive functions.
Left Hemisphere
Essential functions include working memory, concentration, planning, and positive emotion. The main clinical concern is Major Depressive Disorder (MDD).
Right Hemisphere
Critical functions include declarative memory, social awareness, and negative emotions. Principal clinical concerns include Generalized Anxiety Disorder (GAD), fear, and impaired executive functioning.
Frontal lobe damage may result in impaired flexibility and problem solving, increased risk-taking, changes in social behavior, an inability to use external cues, and deficits in emotional self-regulation. Graphic © ART-ur/Shutterstock.com.
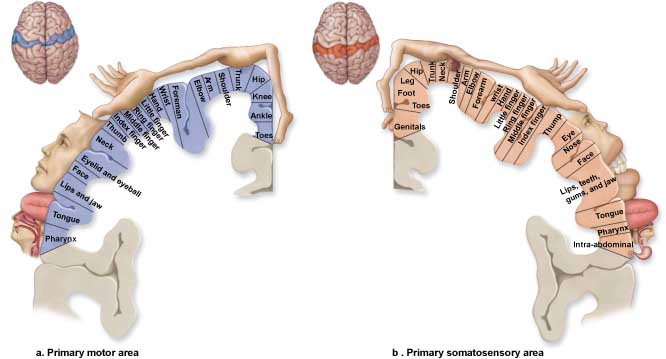
The primary motor cortex is located in the precentral gyrus (Brodmann area 4, BA 4). It organizes the opposite side of the body's muscles and movements required for fine motor coordination in tasks like writing. Lesions can result in loss of motor control, including rigid paralysis. The graphic below, which shows the motor and sensory homunculi, was retrieved from the nccpbwikiproject.
The motor association cortex (premotor cortex), rostral to the primary motor cortex (BA 6), helps program and execute movements. Think of the motor association cortex as the piano player and the primary motor cortex as the keyboard (Carlson & Birkett, 2017). The primary and motor association cortex collectively appear to map behaviors rather than specific muscles or movements (Breedlove & Watson, 2023).
Broca's area, located in the inferior frontal gyrus (BA 44 and 45) of the dominant hemisphere (F7), is concerned with speech production, grammar, language comprehension, and sequencing (Caplan, 2006). Lesions to Broca's area can produce Broca's aphasia, an inability to produce fluent speech despite relatively intact comprehension. Broca's area receives input from Wernicke's area via the arcuate fasciculus (Breedlove & Watson, 2023). Graphic courtesy of Blausen.com staff "Blausen gallery 2014," Wikiversity Journal of Medicine.
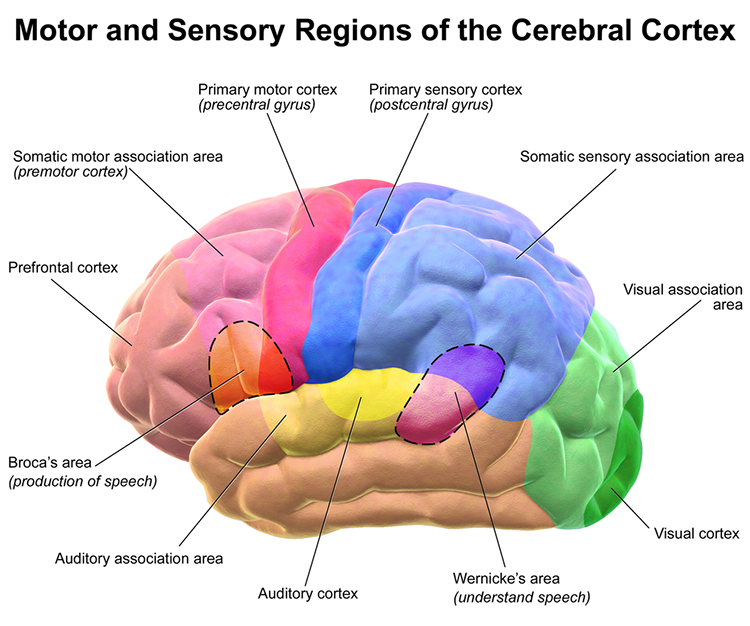
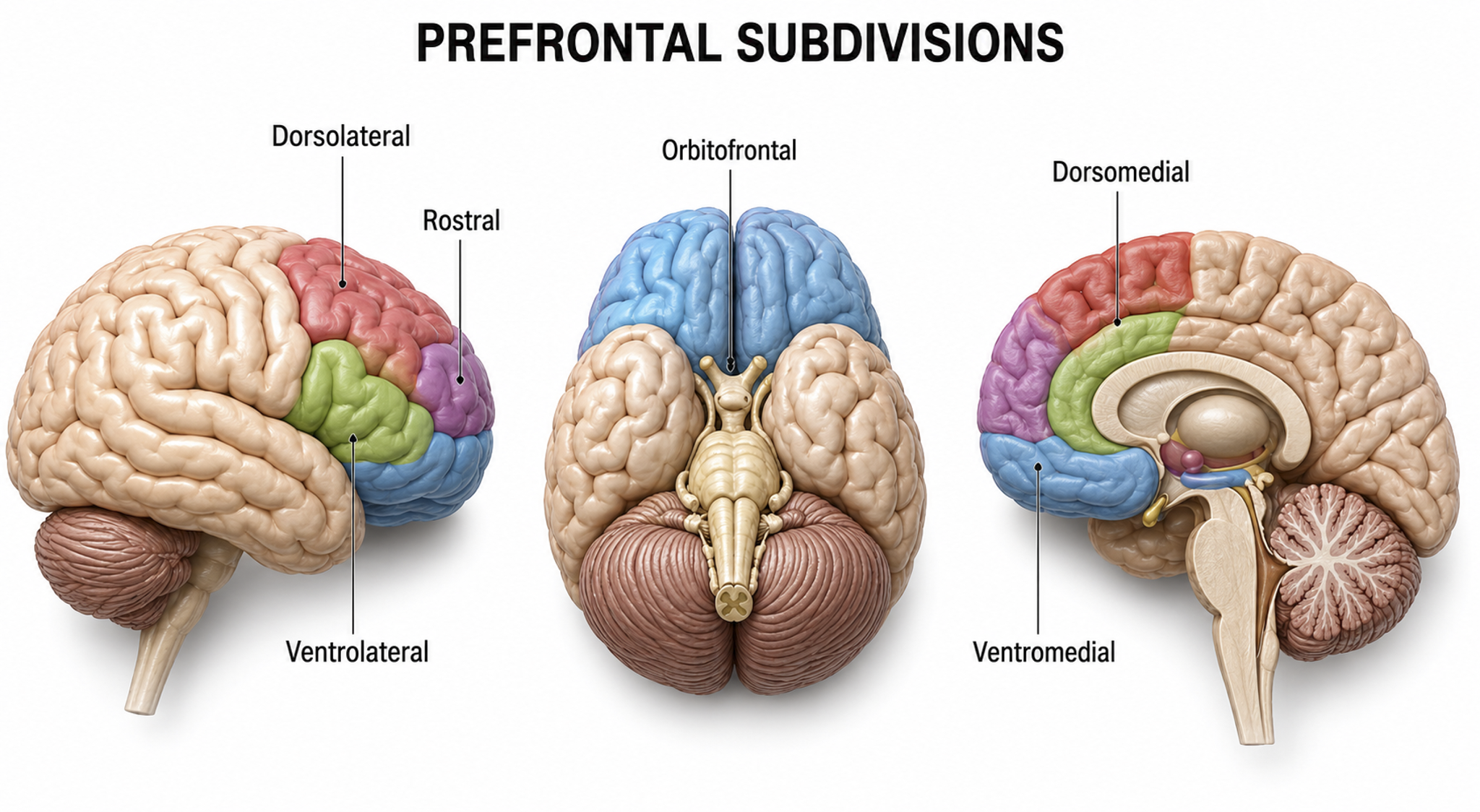
The prefrontal cortex (PFC) (BA 9, 10, 11, 12, 25, 32, 44, 45, 46, 47) is rostral to the motor association area. The PFC participates in executive functions, including attention, working memory, outcome prediction for current and hypothetical actions, goal-directed behavior, problem-solving, planning, and the ability to suppress actions that could lead to unwanted outcomes (Diamond, 2013). The PFC integrates emotion and reward in decision-making (Fuster, 2015).
Important subdivisions of the PFC include the orbitofrontal cortex, ventromedial PFC, and dorsolateral PFC.
The orbitofrontal cortex (OFC) comprises Brodmann areas 10, 11, and 47 in Brodmann's human map, subdivided in the Walker scheme as areas 11, 13, 14, and 47/12 (Kringelbach, 2005); areas 9 and 46 belong to the dorsolateral prefrontal cortex, not the OFC. The OFC aids planning by evaluating the consequences (rewards and punishments) of our actions and helps generate the motivation to ingest drugs. The profound personality changes suffered by Phineas Gage resulted from damage to this subdivision and the ventromedial PFC. The OFC adjusts decision-making based on the stakes involved, enabling us to switch between significant (investments) and trivial (snacks) choices. It compares current options with recent ones, while the anterior cingulate cortex registers our predictions and prediction errors (Kennerley et al., 2011).
The ventromedial prefrontal cortex (VMPFC) corresponds to the ventromedial reward network (Öngür & Price, 2000) and includes BA 10, 25, 32, and parts of 11, and 12. The VMPFC is implicated in decisions where outcomes are uncertain and moral values must be applied to real situations. Patients with VMPFC damage choose options that lead to immediate reward regardless of future cost and do not learn from their mistakes. Since they have difficulty reading social cues, they may not recognize deception, irony, or sarcasm (Zald & Andreotti, 2010) and may struggle to control emotional reactions in social situations, particularly anger and violence (Carlson & Birkett, 2017).
The dorsolateral prefrontal cortex (DLPFC) is located in the middle frontal gyrus and includes BA 9 and 46. The DLPFC shares responsibility with cortical and subcortical networks for executive functions like abstract reasoning, cognitive flexibility, decision-making, inhibition, planning, and working memory (Miller & Cummings, 2007). It exercises the highest cortical level of motor control (Hale & Fiorello, 2004).
The left DLPFC is concerned with approach behavior and positive affect, helping us select positive goals and organize behavior to achieve them. The right DLPFC organizes withdrawal-related behavior and negative affect and mediates threat-related vigilance, playing a role in working memory for object location. In unipolar depression and premenstrual dysphoric disorder, the right DLPFC may be more active than the left, producing the alpha asymmetry pattern that is one of the most commonly targeted findings in neurofeedback for depression.
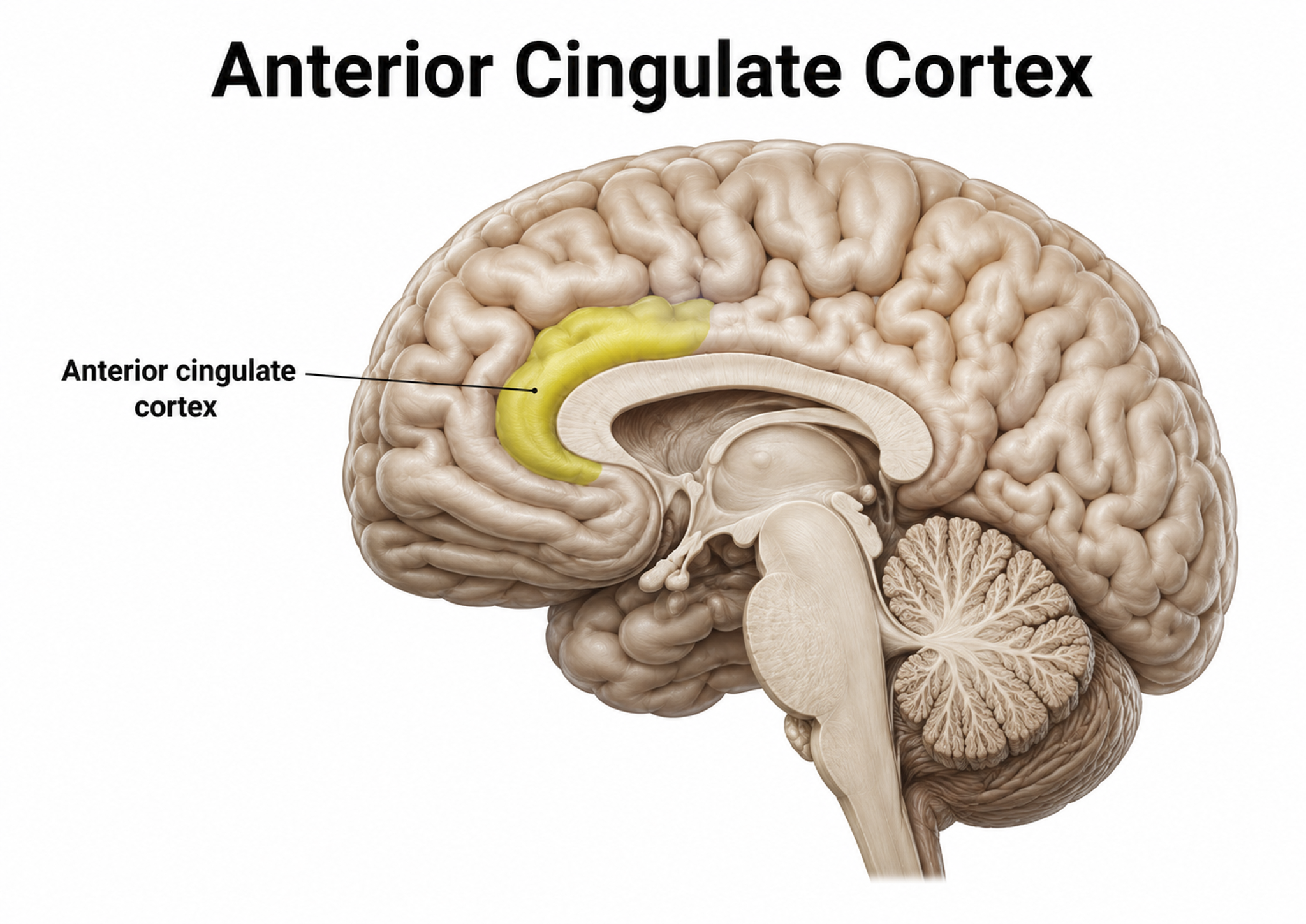
Anterior Cingulate Cortex (ACC)
The cingulate cortex has reciprocal connections with the parahippocampal gyri, integrates limbic functions, and is part of the salience network. Cingulate cortical functions include nurturing, grooming, play, and organizing and managing input/output functions.
The anterior cingulate cortex (ACC) (Fpz and Fz) lies above the corpus callosum (BA 24, 32, 33). The dorsal ACC connects to both the PFC and parietal cortex. The ACC plays a vital role in attention, is activated during working memory, and mediates both emotional and physical pain. It has cognitive (dorsal anterior cingulate) and affective (ventral anterior cingulate) conflict-monitoring components. Graphic courtesy of Geoff B. Hall in Wikimedia Commons.
{kind=link}
The Stroop test illustrates a cognitive monitoring task where color and name information conflict, while discrepancies between facial and vocal cues present an affective conflict. The anterior cingulate recruits other brain areas to resolve these conflicts.

The anterior cingulate gyrus helps us allocate attention: focusing on a target, disengaging, perceiving options, and making adaptive choices. The anterior cingulate gyrus, prefrontal cortex, and caudate function abnormally in children diagnosed with ADHD during selective attention tasks. Importantly, fMRI evaluation showed that neurofeedback could teach children to normalize activity in these structures (Beauregard & Levesque, 2006).
The anterior cingulate gyrus is also involved in motivation and the perception of emotional and physical pain. Eisenberger, Lieberman, and Williams (2003) used fMRI to study brains of subjects who believed that two companions in Cyberball, an online ball-tossing game, had suddenly begun excluding them; their emotional distress activated the anterior cingulate cortex, the same region that evaluates physical pain. deCharms and colleagues (2005) provided real-time fMRI feedback from the rostral anterior cingulate, and subjects learned to increase and decrease its activation, with corresponding increases and decreases in the rated intensity of a noxious thermal stimulus.
Lesions to the cingulate can produce akinetic mutism, a state of markedly reduced spontaneous movement and speech despite preserved wakefulness, in which orienting responses are absent or greatly diminished. Cingulate malfunction can also result in addictive behaviors (alcohol or drug abuse, eating disorders, chronic pain), obsessive-compulsive disorder and OCD spectrum disorders, and "road rage."
Parahippocampal Gyri
The parahippocampal gyri are located within the medial temporal lobe. They form spatial and nonspatial contextual associations, which serve as building blocks for contextual processing, episodic memory, navigation, and scene processing (Aminoff, Kveraga, & Bar, 2013). They may also play a role in emotional responsiveness. Polygon data were generated by Database Center for Life Science (DBCLS). Creative Commons Attribution-Share Alike 2.1 jp.
Parietal Lobes
The parietal lobes (Pz, P3, P4) are posterior to the frontal lobes and are divided into the primary somatosensory cortex and secondary somatosensory cortex. Their main function is to process somatosensory information like pain and touch.
Left Hemisphere
Major functions include attention, association, complex grammar, math, object names, problem-solving, and somatosensation.
Right Hemisphere
Major functions include body boundary awareness, geometry, guiding reaching with the hands, somatosensation, spatial awareness, and spatial perception (Demos, 2019).
The primary somatosensory cortex (S1) is located in the parietal lobe's postcentral gyrus, posterior to the central sulcus (BA 3, 1, and 2). S1 maps touch and pain information from the opposite side of the body. The secondary somatosensory cortex (S2), which lies in the parietal operculum (BA 40 and 43), receives projections from it and maps touch and pain from both sides of the body (Breedlove & Watson, 2023). Graphic adapted from Paskari from Wikimedia Commons.
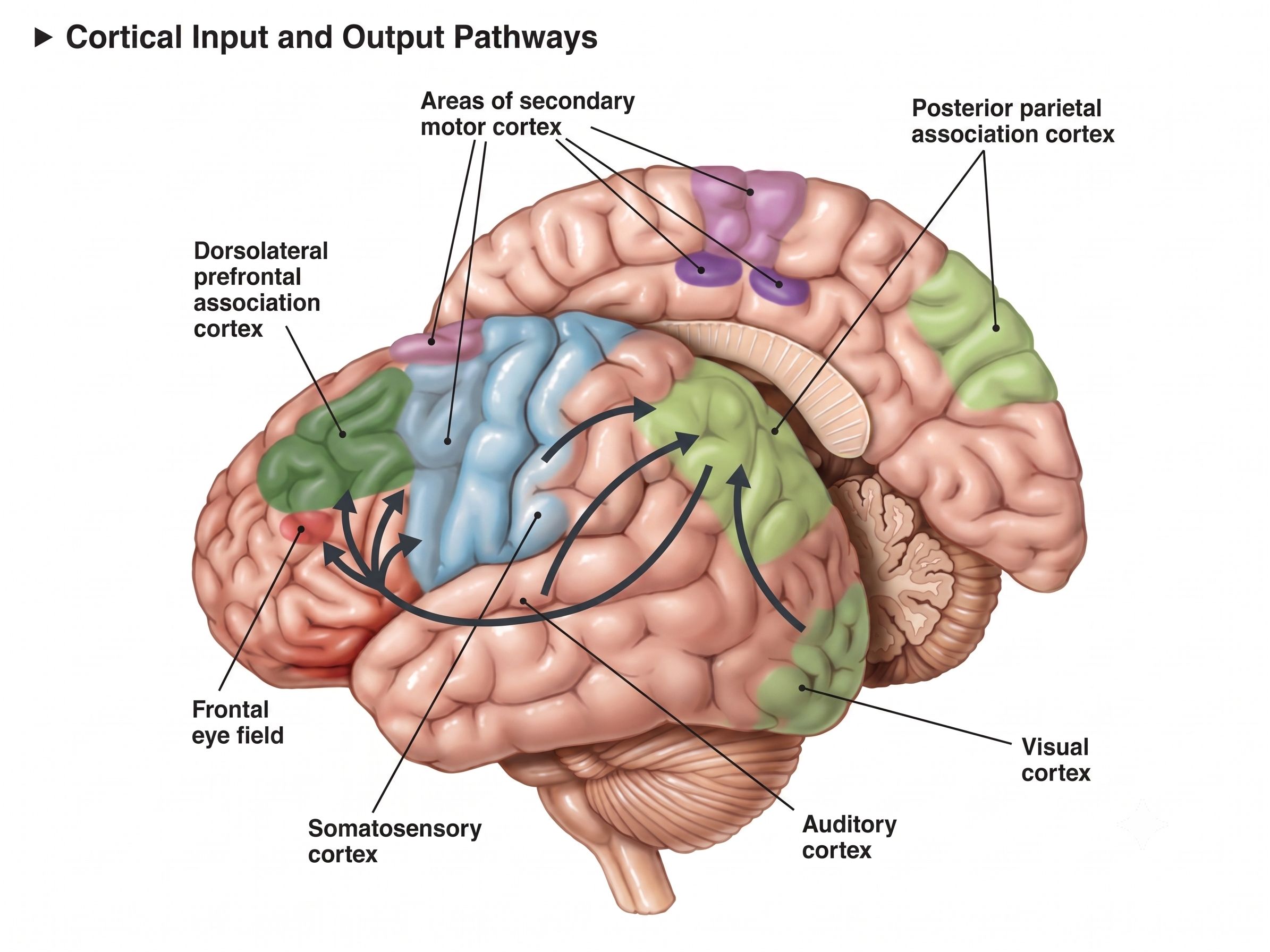
The parietal cortex monitors our preparation for a movement and is responsible for our subjective feeling of intending to move (Sirigu et al., 2004).
The angular gyrus, located near the superior temporal lobe (BA 39), is involved in reading, math, and copying writing.
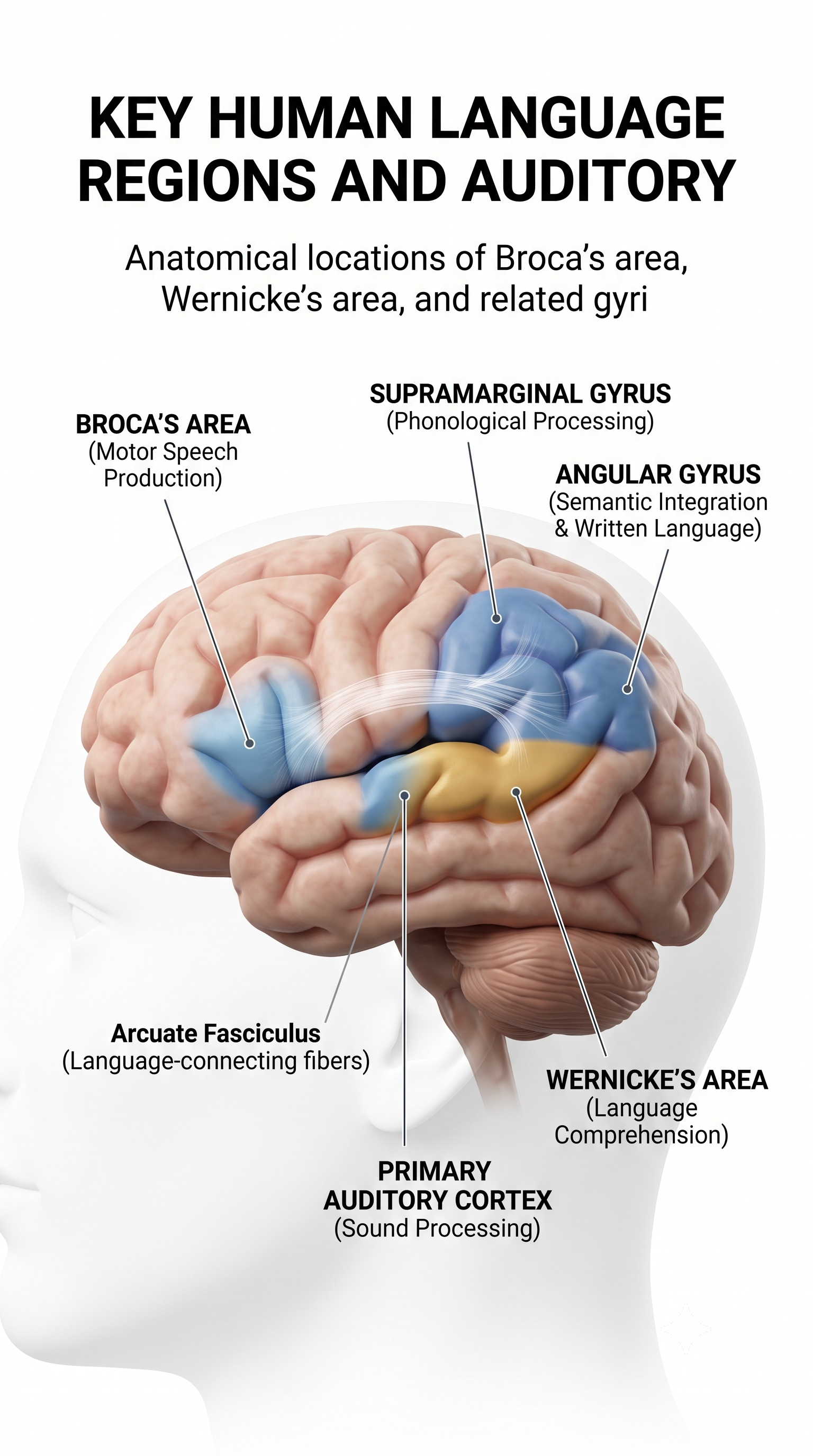
The supramarginal gyrus (BA 40) forms part of the inferior parietal lobule and, like the neighboring angular gyrus (BA 39), functions as a convergence zone where somatosensory, auditory, and visual information are integrated. Positioned at the posterior end of the lateral sulcus, it draws on this multimodal input to support several higher-order functions.
In written language, it contributes to phonological processing during reading and writing, helping map the sounds of speech onto their corresponding letters and words. Its role in verbal working memory reflects its involvement in the phonological store, the component of the phonological loop that briefly holds speech-based information for rehearsal. The right supramarginal gyrus also supports social cognition by helping a person distinguish their own emotional state from that of others, a process central to empathy.
Both the angular gyrus and the supramarginal gyrus are in the vicinity of the P3 electrode site.
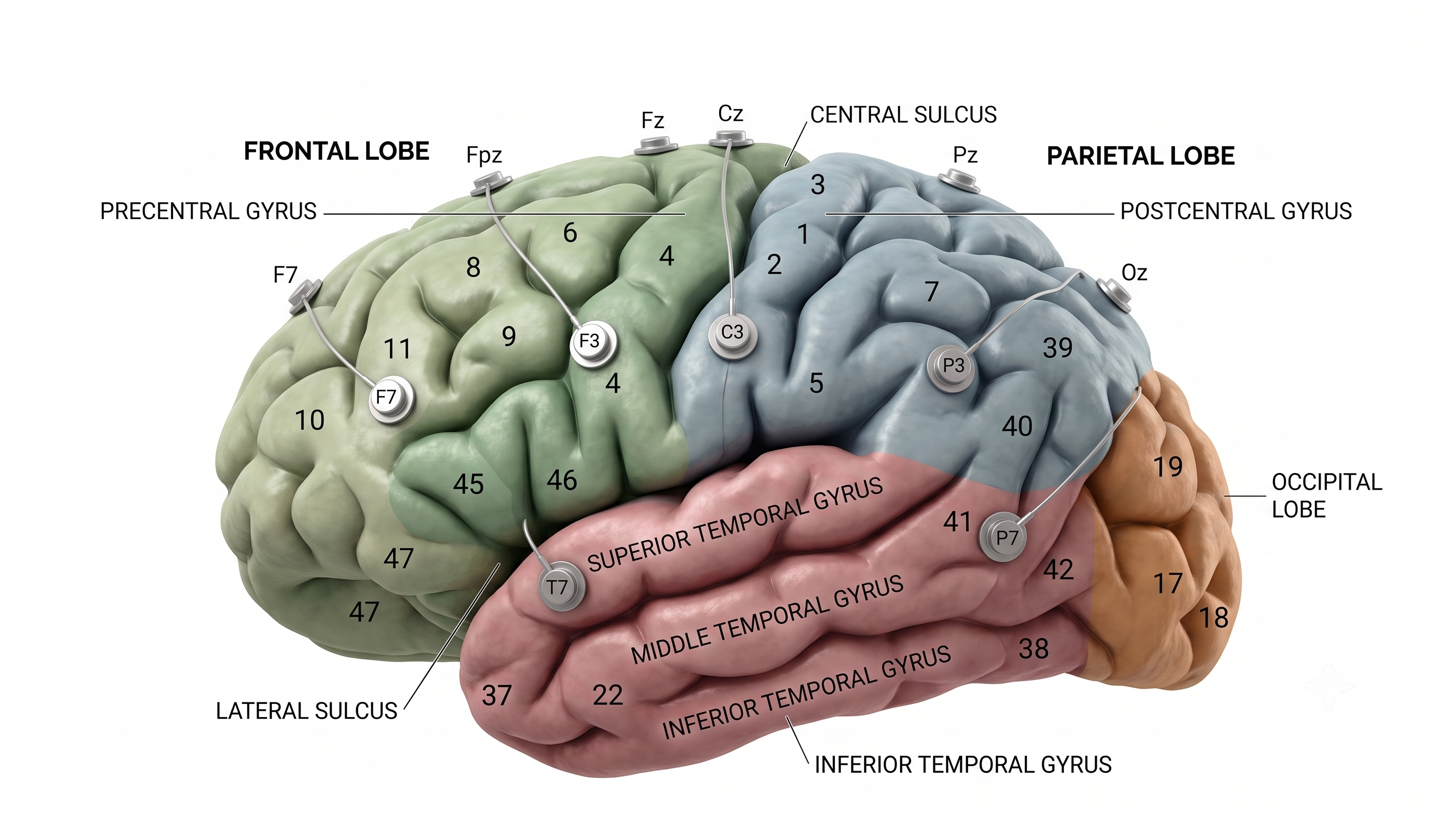
Temporal Lobes
The temporal lobes lie beneath the temporal bone at the sides of the head, below the T3 and T5 electrode sites on the left and the T4 and T6 sites on the right. They contain numerous Brodmann areas, both laterally (20, 21, 22, 37, 38, 41, and 42) and medially (20, 27, 28, 34, 35, 36, 37, and 38); note that several of these areas wrap around from the lateral to the medial cortical surface. The temporal lobes process hearing, smell, and taste information and help us understand spoken language and recognize visual objects and faces (Breedlove & Watson, 2023).
Their medial cortex is involved in emotional experience and new memory formation. In the language-dominant hemisphere (usually the left), Wernicke's area lies beneath electrode sites C3, T3, and T5 and encompasses posterior regions of Brodmann areas 22, 41, and 42.
Wernicke's area, located in the temporoparietal cortex (BA 22) of the dominant hemisphere, is specialized for the comprehension of spoken and written language. Damage can result in an inability to understand the meaning of speech and to construct meaningful sentences, a condition known as Wernicke's aphasia, in which speech output remains fluent but is empty of meaning and carries paraphasias and neologisms. Graphic adapted from Blausen.com in the Wikiversity Journal of Medicine.
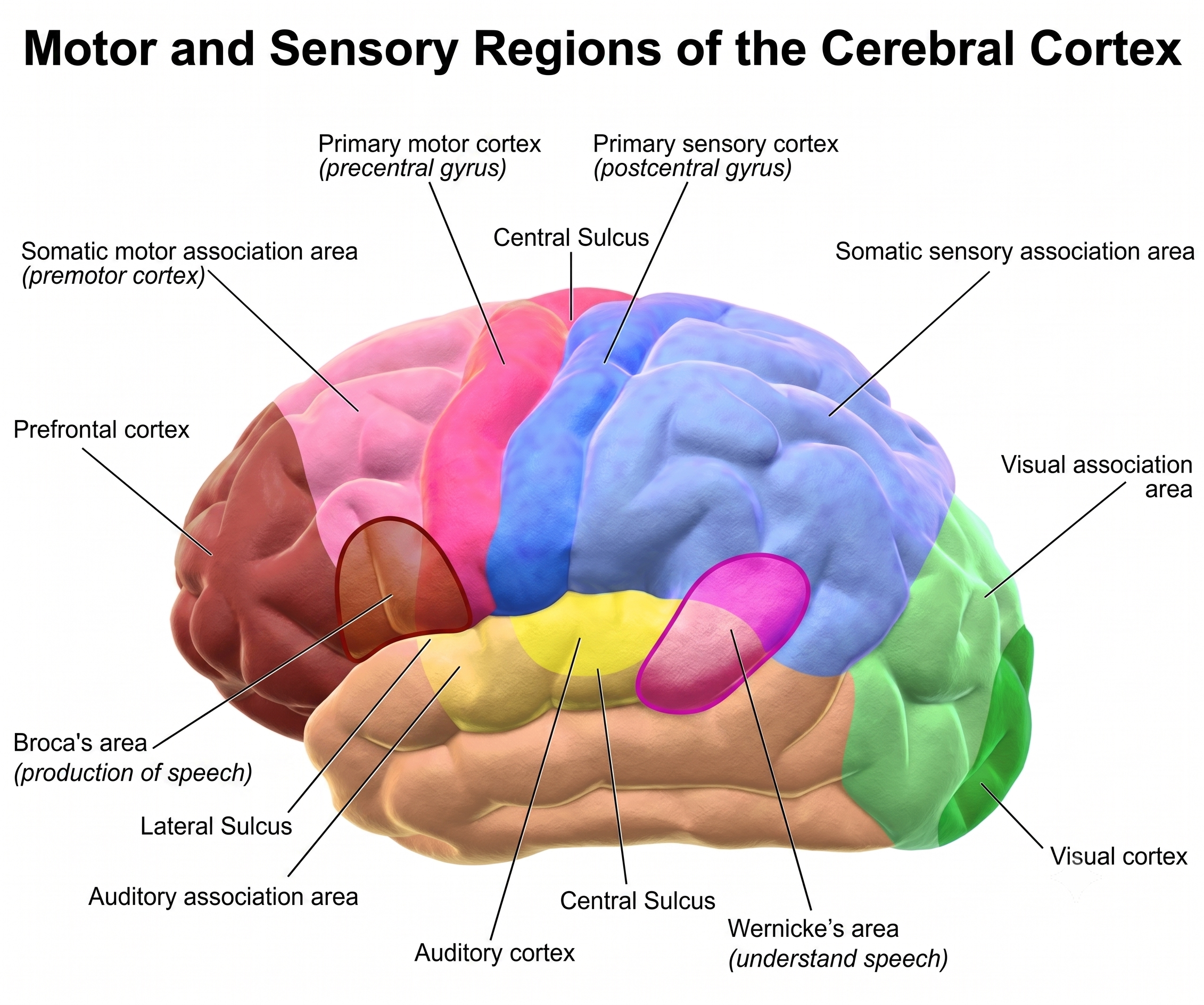
Left Hemisphere
Major functions include affect, declarative memories, language comprehension, perception of movement, reading, and word recognition.
Right Hemisphere
Important functions include face and object recognition, music, and social cues (Demos, 2019).
Occipital Lobes
The occipital lobes (Oz, O1, O2) are posterior to the parietal lobes. The primary visual cortex (V1) is located within the calcarine sulcus (BA 17). The occipital lobes process visual information from the eyes in collaboration with the frontal, parietal, and temporal lobes. Graphic © ART-ur/Shutterstock.com.
Their primary functions are visual, including the analysis of orientation, color, spatial frequency, illusory contours, and complex patterns like concentric and radial stimuli (Breedlove & Watson, 2023). The occipital lobes are the primary source of the posterior dominant alpha rhythm, making them a key region of interest in neurofeedback assessment.
Insular Cortex
The insular cortex lies buried deep within the lateral sulcus, concealed by the frontal, parietal, and temporal opercula, and corresponds to Brodmann area 13. It supports interoception (awareness of internal bodily states), autonomic regulation, emotional processing, and social cognition, and serves as a core hub of the salience network. It detects salient events via afferent pathways and switches between other large-scale networks when such events are identified, affecting attention and working memory. The anterior and posterior insulae interact to regulate autonomic responses to salient stimuli, and interactive communication between the insula and anterior cingulate cortex facilitates motor control (Menon & Uddin, 2010). The right insula mediates awareness of our body, empathy, and understanding others' points of view (Khazan, 2019).
Increased heart rate variability strengthens the connectivity between the ACC and the insula for empathy and the ability to understand others' emotions, feel gratitude, socially connect, understand our own psychophysiological states, and restore nervous system balance. Mindfulness meditation increases insula gray matter and activation. This finding is clinically significant because it suggests that HRV biofeedback and mindfulness-based interventions may produce measurable structural changes in a brain region central to self-awareness and emotional regulation.
The insula functions as an integrative and organizational hub for the salience network, integrating interoceptive awareness, emotional experience, and external perception to facilitate our global perception of the world and our relationship with it. The insula directs specific networks in the processing of salient stimuli and in generating appropriate behavioral responses (Wiebking & Northoff, 2014).
The insula is the primary taste cortex and is activated when you see something that disgusts you or see another person's expression of disgust. Pictures of lovers also activate the anterior insula, as opposed to friends. In neuroeconomic studies, anterior insula activation predicted risk-avoidant financial strategies (choosing bonds instead of stocks), and in the Prisoner's Dilemma game, mutually cooperative decisions also activated this region.
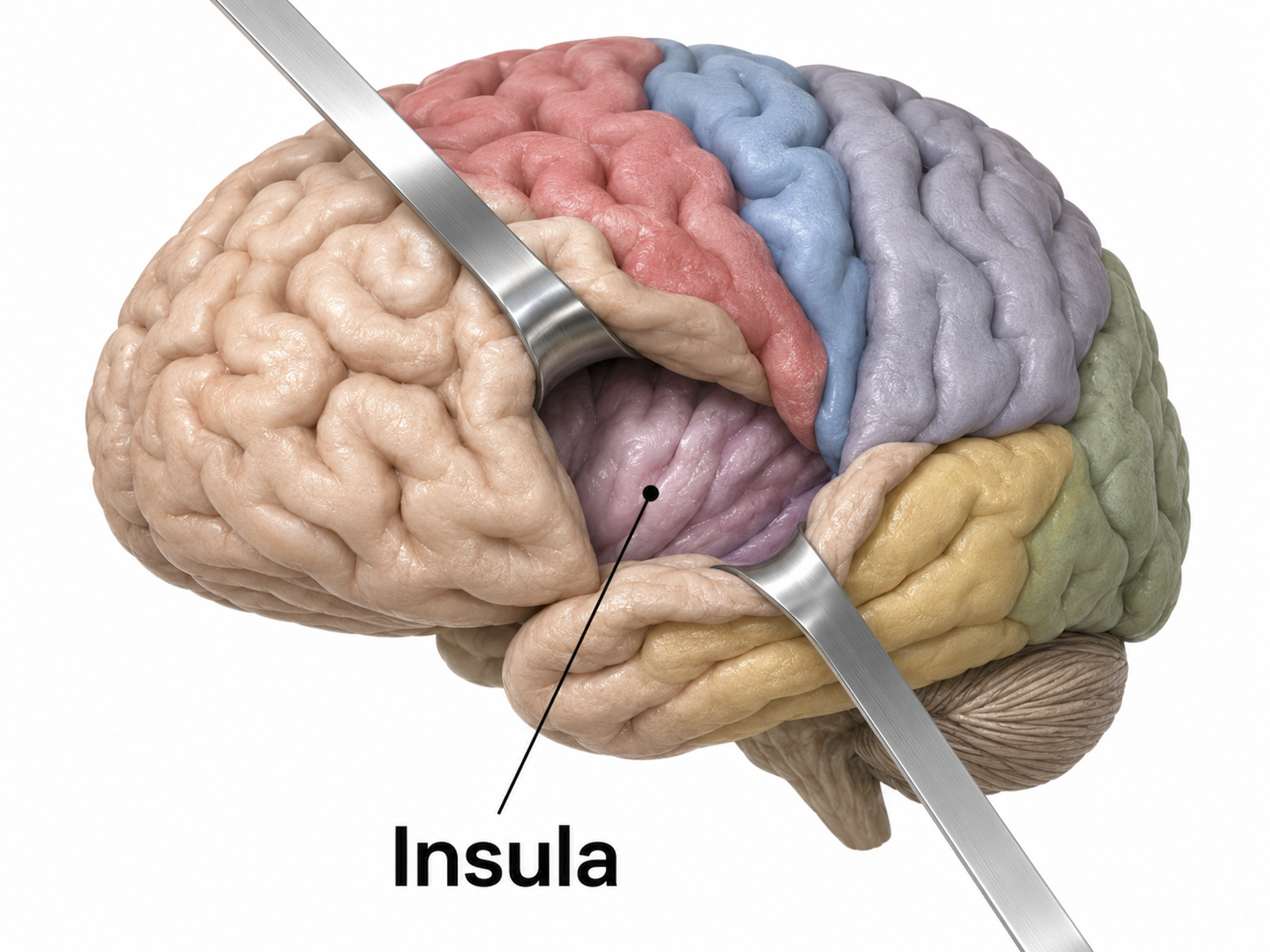
Antonio Damasio has proposed that this region helps map visceral states associated with emotional experience and generate conscious feelings, providing the basis for somatic markers, gut feelings like the discomfort produced by a risky decision.
The insular cortex has been implicated in the experience of pain and basic emotions including anger, disgust, fear, happiness, and sadness. It receives reports of internal states like hunger and drug craving, motivating individuals to engage in consummatory behavior. The insular cortex plays a crucial role in craving and impulse control. It is stimulated by drug-related cues and may activate memories of pleasurable drug-related experiences. Remarkably, smokers whose strokes damaged the insular cortex have been found far more likely than smokers with damage elsewhere to quit easily, immediately, and without relapse, although not all of them quit.
National Institute of Drug Abuse graphic retrieved from Wikimedia Commons and adapted.
Cortical and Subcortical Connections
Neocortical zones are connected with cortical and subcortical regions by specialized fiber tracts: association bundles, projection fibers, and commissural bundles. A meta-networking model proposes that the dynamic interaction of "distributed but relatively specialized networks" mediates brain functions like language (Herbet & Duffau, 2020, p. 1181).
We adapted the figure below from Do et al. (2025), which shows major white matter tracts of the brain.
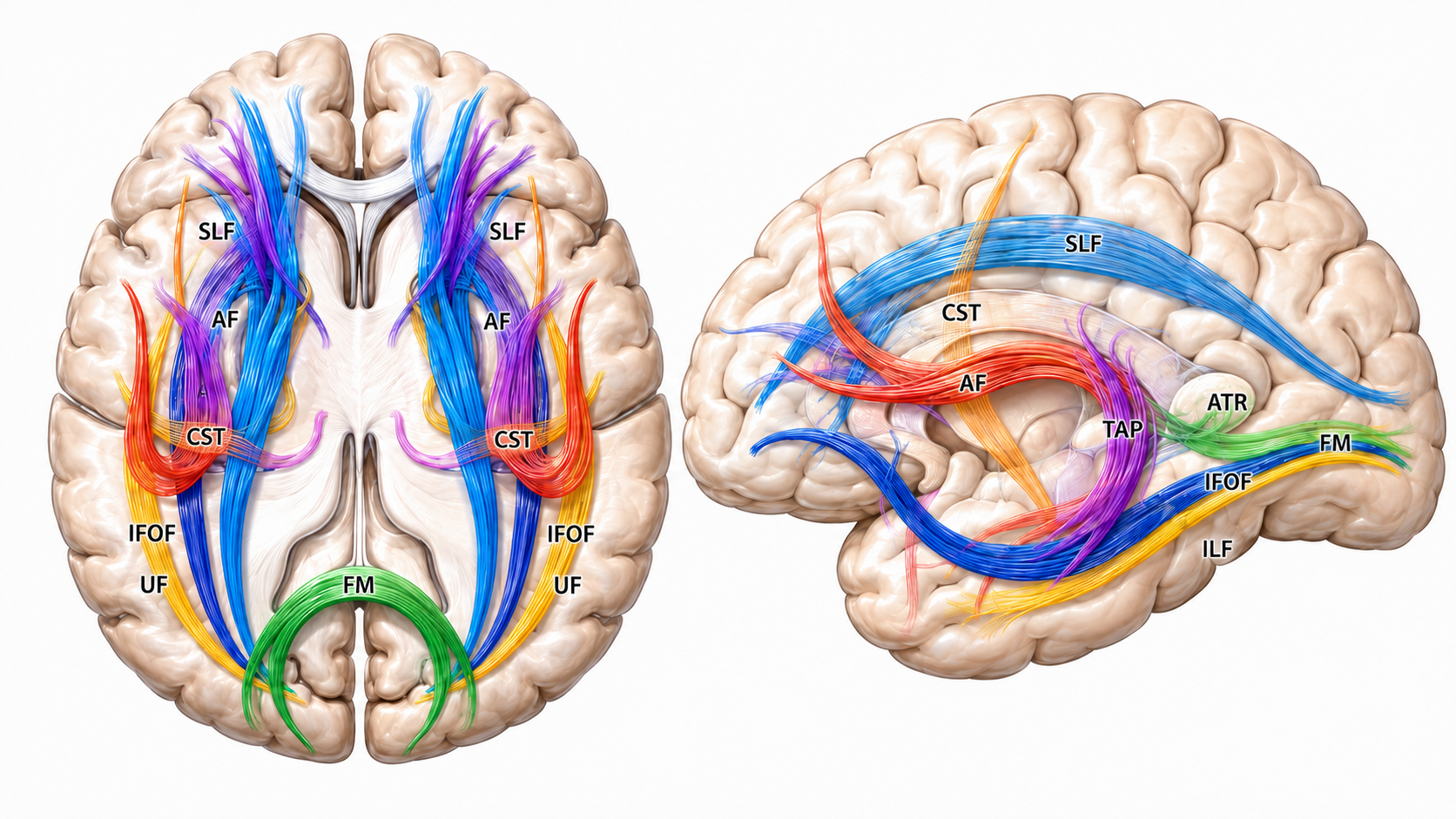
Artistic view of the main white matter tracts in axial view (left panel) and sagittal view (right panel): AF arcuate fasciculus, ATR anterior thalamic radiations, CST corticospinal tract, FM forceps major, IFOF inferior fronto-occipital fasciculus, ILF inferior longitudinal fasciculus, SLF superior longitudinal fasciculus, TAP tapetum, UF uncinate fasciculus. Pencil drawing performed by Nathan Beucler, MD.
Association Bundles
Association bundles link cortical regions located in the same hemisphere. Short association fibers, called U-fibers, arch beneath a sulcus to join adjacent gyri, while long association fasciculi connect widely separated regions. The long fasciculi include the arcuate fasciculus (AF), frontal aslant tract (FAT), inferior fronto-occipital fasciculus (IFOF), inferior longitudinal fascicle (ILF), middle longitudinal fasciculus (MdLF), superior longitudinal fasciculus (SLF), and uncinate fasciculus (UF).
Projection Fibers
Projection fibers connect the cortex with structures deep in the brain, the brainstem, and spinal cord. They include the frontostriatal tract (FST), which connects the premotor cortex with the caudate nucleus and putamen, as well as the thalamocortical, optic, and pyramidal tracts. Tractography animation Alfred Anwander, CC BY-SA 4.0, via Wikimedia Commons.
{kind=link}
Commissural Bundles
The left and right hemispheres communicate using three commissures, or axon tracts. The corpus callosum is the largest tract and connects the left and right frontal, parietal, and occipital lobes. Certain conditions, such as prenatal exposure to alcohol and other drugs, may result in agenesis of the corpus callosum, in which part or all of this fiber bundle is missing. Graphic © decade3d - anatomy online/Shutterstock.com.
The anterior commissure, shown above the third ventricle at the bottom of the diagram, is considerably smaller than the corpus callosum and connects the left and right temporal lobes and the hippocampus and amygdala. Anterior commissure graphic adapted from Winter, T. J. & Franz, E. A., CC BY 3.0, via Wikimedia Commons.
{kind=link}
The posterior commissure, located below the corpus callosum, connects the right and left diencephalon and mesencephalon (Breedlove & Watson, 2023). Corpus callosum graphic © Image Trading Source Ltd/Shutterstock.com.
Subcortical Structures
Subcortical regions, including the thalamus, basal ganglia, and limbic system (e.g., amygdala, hippocampus, hypothalamus), regulate fundamental functions like sensory processing, movement, and emotion. Sometimes the cerebellum is included among subcortical structures. These regions also contribute to the regulation of various qualities of thinking and emotion.
Thalamus
The thalamus consists of specialized nuclei that process and relay data to and from the telencephalon (cerebral cortex, basal ganglia, and limbic system). The thalamus analyzes all sensory data except olfaction before distributing this information to the cortex via thalamocortical afferent fibers (Breedlove & Watson, 2023). The cortex sends information back to the thalamus via corticothalamic fibers to adjust its processing, creating feedback loops that are crucial for generating several EEG rhythms. The thalamus helps regulate arousal, sleep, and wakefulness (Steriade & Llinás, 1988) and, through its functional connection to the hippocampus, plays a crucial role in episodic memory (Aggleton et al., 2010).
The thalamus contributes to SCPs, delta, theta, alpha, SMR activity, and beta-gamma activity (Thompson & Thompson, 2015). This makes the thalamus the single most important subcortical structure for understanding EEG generation. Thalamus graphic © decade3d - anatomy online/Shutterstock.com.
Basal Ganglia
The basal ganglia (comprising the caudate nucleus, putamen, globus pallidus, subthalamic nucleus, and substantia nigra) modulate movement. The basal ganglia, prefrontal cortex, cingulate cortex, and parietal cortex are involved in self-awareness, attention, and emotional regulation. Researchers have identified several distinct prefrontal-basal ganglia-thalamic loops that are implicated in a range of psychiatric conditions. Graphic © Kateryna Kon/Shutterstock.com.
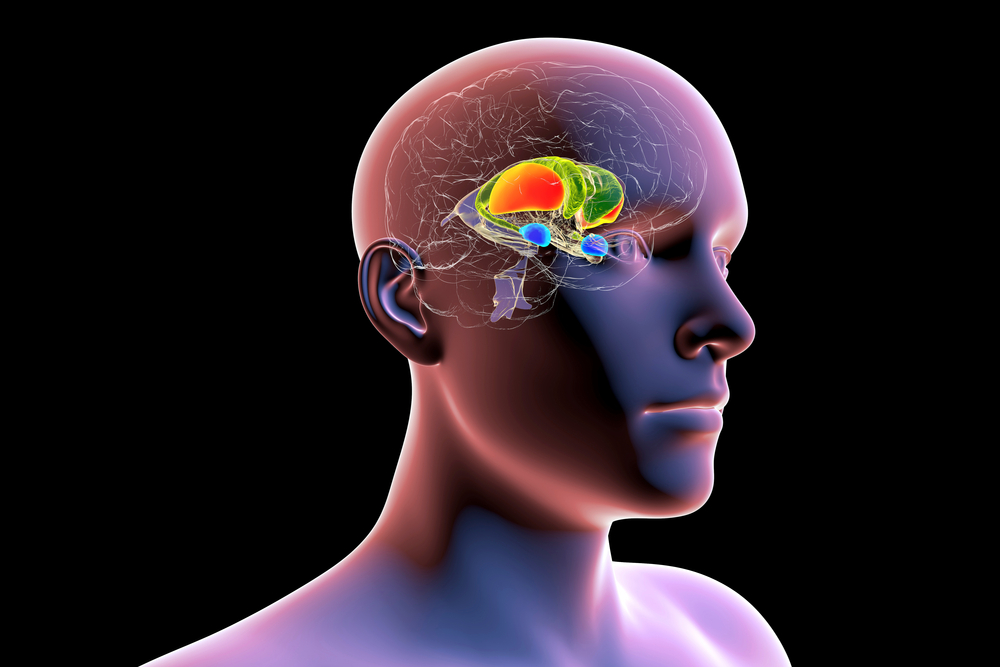
Limbic System
The limbic system is a widespread network of nuclei involved in emotion, motivation, learning, memory, and navigation. Three important limbic structures are the hippocampus, amygdala, and septal nuclei, along with the hypothalamus, anterior thalamus, and cingulate gyrus (Breedlove & Watson, 2023). Limbic system graphic © decade3d - anatomy online/Shutterstock.com.
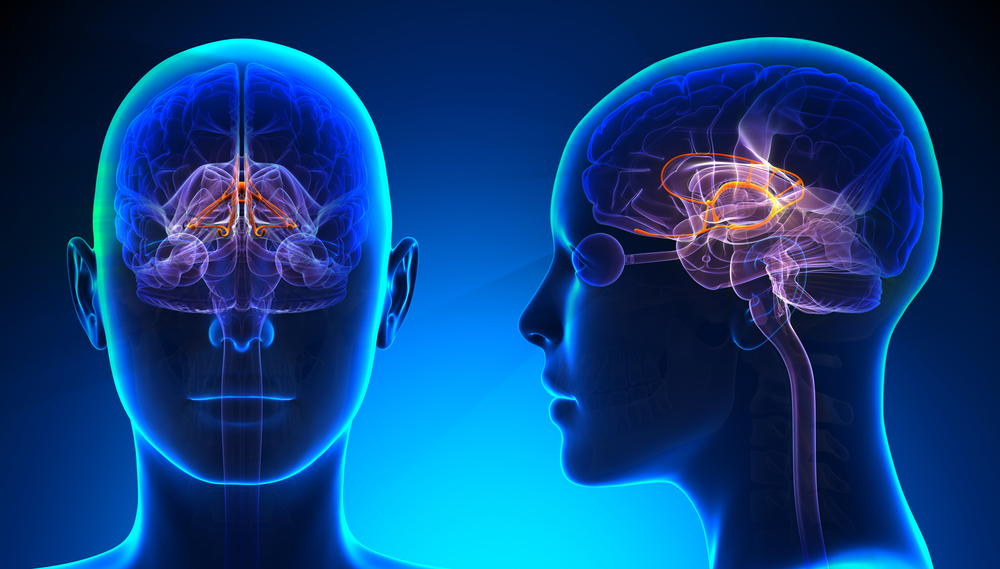
Hippocampus
The hippocampus is a seahorse-shaped limbic structure required to form declarative memories. It plays a vital role in emotion, navigation, spatial memory, and dampening the endocrine stress response. The hippocampus simultaneously integrates emotional, auditory, and visuospatial information to create episodic memories and contains leukocyte receptors, making it part of the feedback loop for immune system regulation. Hippocampal neurons and networks that include it are important sources of the theta rhythm (Amzica & Lopes da Silva, 2018).
Hippocampal Brainwaves Travel in Two Directions
The old-school view held that hippocampal brainwaves travel in one direction, but this model could not explain how the hippocampus integrates information from various interconnected specialized systems. The new-school view, based on recording from human participants undergoing brain surgery, is that brainwaves travel through the hippocampus in both directions: from back to front and from front to back (Kleen et al., 2021). Cognitive activity differentially influences the direction of movement for low (e.g., 1.9 Hz) and high (e.g., 13.8 Hz) frequency waveforms.
Human hippocampal neuron graphic © Kateryna Kon/Shutterstock.com.
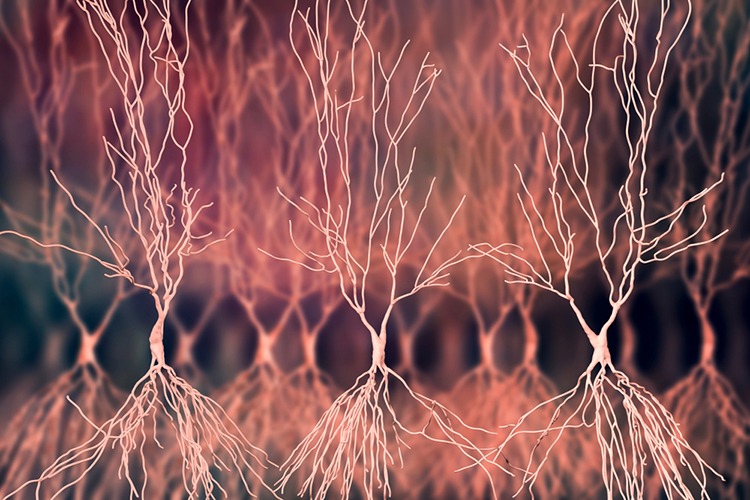
Watch Sam Kean's TED-Ed Talk, What Happens When You Remove the Hippocampus. Hippocampus graphic © decade3d - anatomy online/Shutterstock.com.
Amygdala
The amygdala is a collection of nuclei located deep within the medial temporal lobes at the end of the hippocampus. It is an essential limbic structure that includes, among others, the lateral nucleus and the central nucleus.
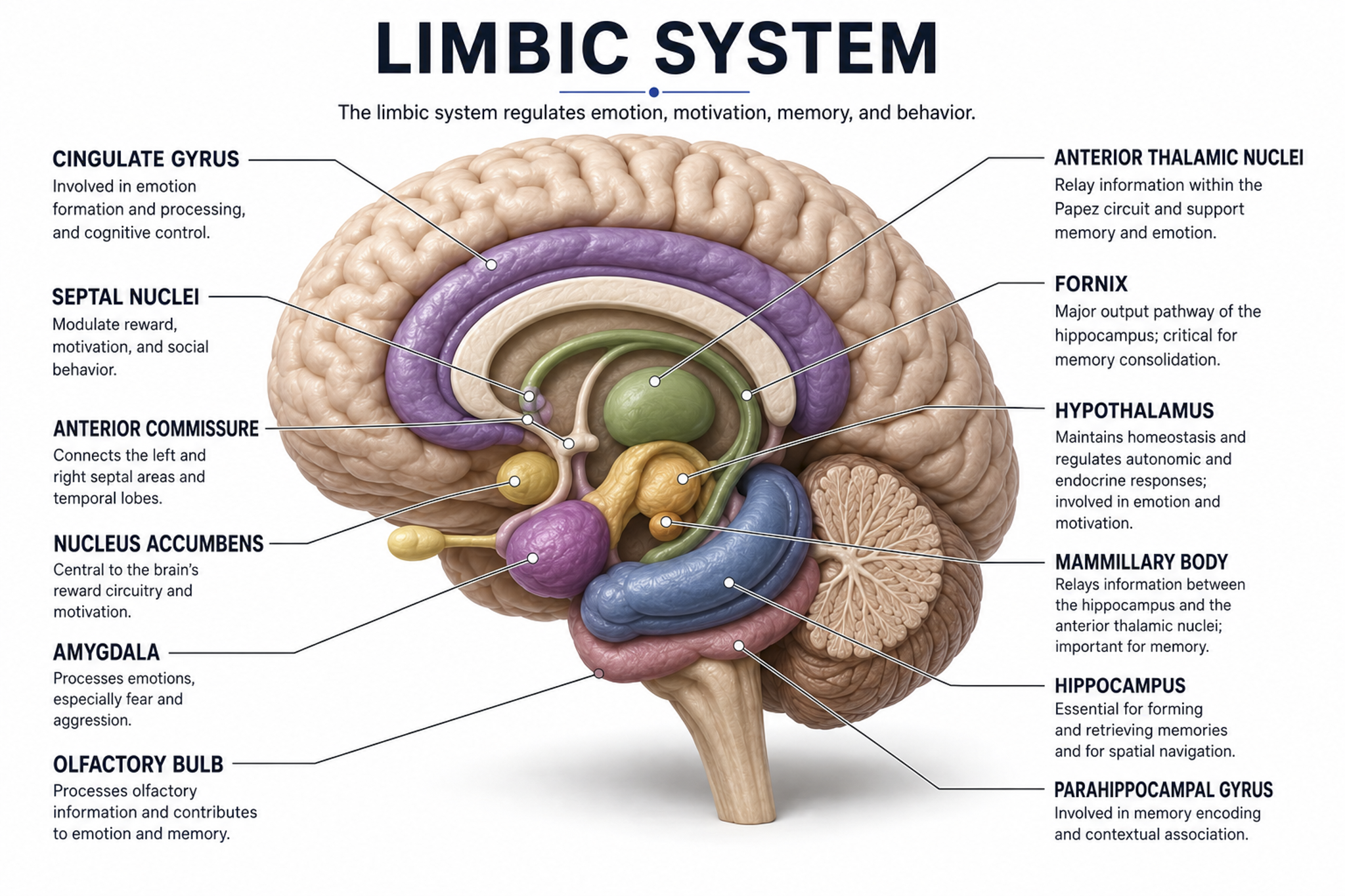
The lateral nucleus processes sensory information and distributes it throughout the amygdala. The central nucleus orchestrates the nervous system's response to important stimuli by activating circuits in the brainstem (autonomic arousal), basal ganglia, and periaqueductal gray (defensive behavior). The amygdala plays a crucial role in learning about the consequences of our actions and creating declarative memories for events with emotional significance (Breedlove & Watson, 2023). Understanding the amygdala's role in fear conditioning is particularly relevant for clinicians treating PTSD and anxiety disorders. Amygdala graphic © decade3d - anatomy online/Shutterstock.com.
Septal Nuclei
The septal nuclei are a limbic structure containing several nuclei involved in emotion, control of aggressive behavior, reward, and addiction (Breedlove & Watson, 2023). The septohippocampal system contributes to the theta rhythm (Amzica & Lopes da Silva, 2018). Septal nuclei graphic © MattL_Images/Shutterstock.com.
In summary, the cortical lobes and subcortical structures work together through extensive reciprocal connections to support cognition, emotion, movement, and memory. For biofeedback and neurofeedback practitioners, recognizing which structures contribute to specific EEG patterns, and which clinical disorders involve dysfunction in these regions, is the foundation of effective assessment and protocol design.
Clinical Application
Excess left frontal alpha, part of the alpha asymmetry pattern, is one of the most commonly targeted findings in neurofeedback for depression. In unipolar depression, the right dorsolateral prefrontal cortex may be more active than the left, and higher alpha over the left frontal region reflects reduced approach-related activation. A clinician who understands this structure and function relationship can design an asymmetry protocol rather than simply chasing a single amplitude value. Interpreting the finding in the context of the client's history and symptoms remains essential.
Key Takeaways
The four cortical lobes and the insula have distinct roles: the frontal lobe governs executive function, movement, and speech; the parietal lobe processes somatosensation and spatial information; the temporal lobe handles hearing, language, and memory; and the occipital lobe processes vision and is the main source of the posterior alpha rhythm. The prefrontal subdivisions differ in function, with the orbitofrontal cortex weighing rewards and punishments, the ventromedial prefrontal cortex applying values to uncertain decisions, and the dorsolateral prefrontal cortex directing executive control. Left and right dorsolateral prefrontal cortex support approach with positive affect versus withdrawal with negative affect, an imbalance reflected in the alpha asymmetry targeted in depression. The insula anchors the salience network for interoception, and higher heart rate variability strengthens its connectivity with the anterior cingulate cortex. Subcortical structures, especially the thalamus, basal ganglia, and limbic system, generate and modulate EEG rhythms and support movement, memory, and emotion.
Check Your Understanding
- Match each lobe to a primary function: frontal, parietal, temporal, and occipital.
- Distinguish the roles of the orbitofrontal cortex, the ventromedial prefrontal cortex, and the dorsolateral prefrontal cortex.
- How do the left and right dorsolateral prefrontal cortex differ in the affect and behavior they support, and how does this relate to the alpha asymmetry seen in depression?
- What network does the insula anchor, and how does heart rate variability relate to insula connectivity?
- Which subcortical structure is the dominant pacemaker for EEG rhythms, and which limbic structures contribute to the theta rhythm?
Brodmann Areas
This section covers the Brodmann classification system, its historical foundations, and its clinical relevance to biofeedback and neurofeedback practice. We will examine how specific Brodmann areas are implicated in disorders you will encounter in clinical settings, from ADHD and depression to PTSD and schizophrenia.
Historical Foundations
Brodmann areas are regions of the cerebral cortex defined by their distinct cytoarchitectonic characteristics (the organization, density, and layering of neurons), first delineated by German neurologist Korbinian Brodmann in the early twentieth century. Using Nissl staining techniques, Brodmann examined cortical cell organization across different brain regions in multiple mammalian species, including humans.
Brodmann compared the human cortex with that of a range of other mammals, including primates, rodents, and marsupials, publishing maps for humans and eight other species and enabling comparative conclusions about cortical organization (Garey, 1994). This comprehensive approach produced a map whose areas are numbered 1 through 52, each characterized by unique cytoarchitectonic features. Roughly 43 to 47 of those areas were delineated in the human cortex; the remaining numbers were defined only in non-human species, which is why numbers such as 12 through 16 and 48 through 51 are absent from most human maps. His work laid the foundation for modern functional neuroanatomy by linking structural variations in the cortex to specific sensory, motor, and cognitive functions (Brodmann, 1909). Biofeedback practitioners continue to use these divisions when interpreting qEEG findings and designing neurofeedback protocols. Brodmann graphic © sciencepics/Shutterstock.com.
This figure superimposes the 10-20 System electrode sites on the Brodmann areas of the brain's lateral convexity.
The Functional Significance of Specific Brodmann Areas
The Brodmann map remains a widely used anatomical reference in both clinical and research neuroanatomy. For example, Brodmann area 4 corresponds to the primary motor cortex in the precentral gyrus and is critical for voluntary motor control (Penfield & Boldrey, 1937). Area 17, the primary visual cortex in the occipital lobe, is the initial cortical recipient of visual input from the lateral geniculate nucleus (Zeki, 1993). The use of Brodmann areas facilitates the identification of functional zones during neurosurgical procedures and functional imaging studies, aiding in the interpretation of brain activity and lesions.
Language Processing and Brodmann Areas
Language processing illustrates how Brodmann areas map onto clinical function. Broca's area (BA 44 and 45) in the dominant hemisphere's inferior frontal gyrus supports speech production and grammar, while Wernicke's area (BA 22) in the temporal cortex supports speech perception and comprehension. Damage to either area produces distinct aphasia syndromes that clinicians can identify through neuropsychological testing and correlate with qEEG findings. The arcuate fasciculus connecting these regions is a critical white matter tract whose integrity affects language network coherence.
Neuroimaging and Cortical Mapping
Recent advancements in neuroimaging techniques, such as fMRI and PET, have enabled researchers to refine the functional mapping of Brodmann areas with greater precision. Probabilistic cytoarchitectonic atlases, such as those developed by the Jülich Brain Atlas project, have supplemented Brodmann's static classification with data derived from multiple brains, allowing for population-level variability in cortical organization (Eickhoff et al., 2005). This evolution reflects the growing recognition that while Brodmann's map provides a valuable framework, individual differences in cortical morphology and function necessitate more dynamic and data-driven mapping approaches.
Relevance and Limitations
Brodmann areas continue to serve as a foundational framework for understanding cortical organization and function in both clinical and research contexts, from neurosurgical planning to the study of cognitive processes and neuropsychiatric disorders. However, ongoing advances in neuroimaging and connectomics are gradually supplementing this early anatomical schema with more detailed and individualized models of brain organization. For biofeedback practitioners, Brodmann areas remain the standard language for communicating about cortical regions, but clinicians should recognize that individual brains vary and that a given Brodmann area may not occupy the exact same cortical territory in every client.
Researchers have revised the Brodmann maps and correlated areas with their functions. The Brodmann maps below were contributed by Mark Dow, Research Assistant at the Brain Development Lab, the University of Oregon to Wikimedia Commons.
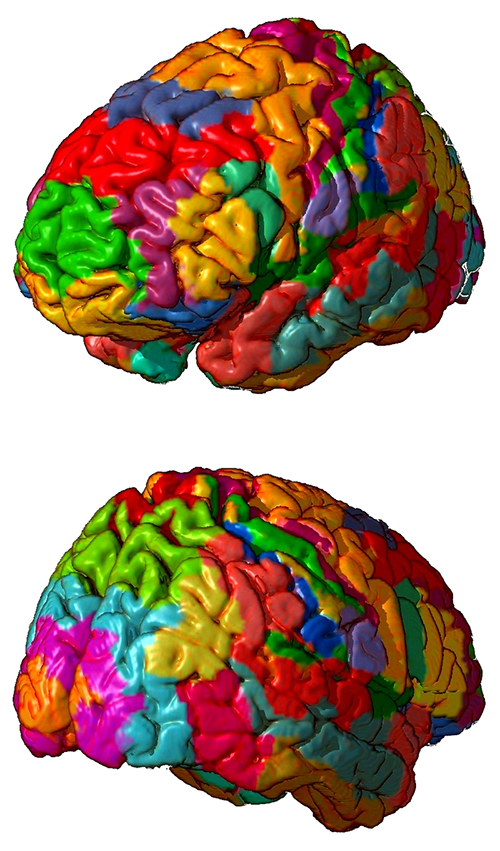
Areas 3, 1, and 2: Primary Somatosensory Cortex (S1)
The primary somatosensory cortex (S1) is a critical region for processing somatosensory information in the brain. It is involved in processing touch, proprioception, and temperature. These four cortical areas contain separate somatotopic maps (Purves, 2018). Graphic © Big8/Shutterstock.com..jpg)
Brodmann areas
The S1 is located in the postcentral gyrus, mainly in Brodmann areas 3, 1, and 2. These areas have distinct functions; Brodmann area 3 receives and processes cutaneous and proprioceptive inputs, area 1 processes tactile stimuli, and area 2 integrates proprioceptive and tactile inputs (Kaas, 2008).Location
The S1 is located in the parietal lobe, immediately posterior to the central sulcus. It is bordered by the primary motor cortex (M1) anteriorly and the secondary somatosensory cortex (S2) posteriorly. The closest sites are C3 and C4, which overlie the central sulcus (Jasper, 1958).Connections
The S1 strongly connects with other cortical and subcortical areas, including the M1, premotor cortex, supplementary motor area, posterior parietal cortex, and thalamus (Lemon, 2008). These connections are essential for sensorimotor integration and control.Participation in brain networks
The S1 is a critical node in the somatosensory network, which includes other areas like the S2, insular cortex, and parietal operculum. It also participates in the sensorimotor network, interacting with the motor and premotor cortices (Sepulcre, 2012).Functions
The S1 is crucial for processing somatosensory information like touch, proprioception, and temperature. It plays a significant role in perceiving object features, body awareness, and sensorimotor integration.Role in clinical disorders
The altered functioning of the S1 has been implicated in various clinical conditions, including neuropathic pain (Baliki et al., 2011), phantom limb pain (Makin et al., 2013), and stroke-related sensory deficits (Carey et al., 2002).Area 4: Primary Motor Cortex (M1)
The primary motor cortex (M1) is a key region in the brain responsible for the execution of voluntary movements. Graphic © Big8/Shutterstock.com.Brodmann areas
The M1 is located in the precentral gyrus, mainly in Brodmann area 4. It contains large pyramidal neurons, known as Betz cells, essential for motor control (Geyer et al., 1996).Location
The M1 is situated in the frontal lobe, immediately anterior to the central sulcus. It is bordered by the posteriorly primary somatosensory cortex (S1) and anteriorly premotor cortex. The closest sites are C3 and C4, which overlie the central sulcus (Jasper, 1958).Connections
The M1 strongly connects with various cortical and subcortical areas, including the S1, premotor cortex, supplementary motor area, posterior parietal cortex, and thalamus (Lemon, 2008). These connections are critical for sensorimotor integration and control.Participation in brain networks
The M1 is a central node in the sensorimotor network, interacting with the somatosensory cortex, premotor cortex, and other motor-related areas (Sepulcre, 2012).Functions
The primary function of the M1 is the execution of voluntary movements. M1 neurons primarily control movements rather than discrete muscles (Breedlove & Watson, 2023). It is critical in planning, controlling, and coordinating complex motor tasks.Role in clinical disorders
Alterations in M1 function have been implicated in various clinical conditions, including motor deficits following stroke (Ward, 2004), Parkinson's disease (Wu & Hallett, 2013), and motor neuron diseases like amyotrophic lateral sclerosis (ALS; Kew & Leigh, 1997).Areas 5 and 7: Somatosensory Association Cortex (SAC)
The somatosensory association cortex (SAC) is involved in the integration and interpretation of somatosensory information coming from the primary somatosensory cortex (S1). Graphic © Big8/Shutterstock.com.Brodmann areas
The SAC is mainly located in Brodmann areas 5 and 7 within the posterior parietal cortex (Culham & Kanwisher, 2001).Location
The SAC is situated in the parietal lobe, superior to the primary somatosensory cortex (S1), and posterior to the postcentral gyrus. The closest sites are likely P3 and P4, which overlie the parietal cortex.Connections
The SAC strong connects with various cortical and subcortical regions, including the S1, primary motor cortex (M1), premotor cortex, supplementary motor area, posterior parietal cortex, and thalamus (Cavada & Goldman-Rakic, 1989). These connections are critical for sensorimotor integration, spatial awareness, and higher-order sensory processing.Participation in brain networks
The SAC is a key node in the somatosensory network, including areas like the S1, S2, and insular cortex. Additionally, it is part of the dorsal attention network, which is involved in attentional control and spatial processing (Corbetta & Shulman, 2002).Functions
The SAC is essential for integrating and interpreting somatosensory information, including tactile and proprioceptive stimuli. It plays a significant role in sensorimotor integration, spatial awareness, and attention.Role in clinical disorders
Alterations in SAC function have been implicated in various clinical conditions, including somatosensory neglect (Vallar et al., 2003), spatial processing deficits (Whitlock et al., 2012), and somatosensory deficits in autism spectrum disorder (Cascio et al., 2012).Area 6: Supplementary Motor Cortex and Premotor Cortex
The supplementary motor cortex (SMA) and premotor cortex (PMC) are critical regions for planning and executing voluntary movements. Graphic © Big8/Shutterstock.com.Brodmann areas
The SMA is primarily located in Brodmann area 6, on the medial aspect of the frontal lobe (Picard & Strick, 2001). The PMC is also predominantly found in Brodmann area 6 but is located on the lateral aspect of the frontal lobe (Wise et al., 1997).Location
The SMA is located in the medial part of the frontal lobe, superior to the cingulate sulcus and anterior to the paracentral lobule. The PMC is situated in the lateral part of the frontal lobe, anterior to the primary motor cortex (M1). The closest sites are likely FC3 and FC4, which overlie the dorsolateral prefrontal cortex.Connections
Both the SMA and PMC have strong connections with various cortical and subcortical areas, including the M1, primary somatosensory cortex (S1), posterior parietal cortex, and basal ganglia (Lemon, 2008; Nachev et al., 2008). These connections are critical for sensorimotor integration, movement planning, and execution.Participation in brain networks
The SMA and PMC are central nodes in the sensorimotor network, interacting with the M1, S1, and other motor-related areas (Sepulcre, 2012).Functions
The SMA and PMC are essential for motor planning, execution, and coordination of complex movements. The SMA is particularly involved in initiating and controlling internally generated movements, while the PMC is more concerned with the planning and executing visually-guided movements (Wise et al., 1997; Picard & Strick, 2001).Role in clinical disorders
Alterations in SMA and PMC function have been implicated in various clinical conditions, including movement disorders like Parkinson's disease (Wu & Hallett, 2013), apraxia (Haaland et al., 2000), and motor deficits following stroke (Ward, 2004).Area 8: Frontal Eye Field (FEF)
The frontal eye field (FEF) is essential for the control of eye movements and visual attention. Graphic © Big8/Shutterstock.com.Brodmann areas
The FEF is predominantly in Brodmann area 8, located in the dorsolateral prefrontal cortex (Paus, 1996).Location
The FEF is located in the anterior bank of the precentral sulcus within the dorsolateral prefrontal cortex, close to the border with the primary motor cortex (M1) (Paus, 1996). The closest sites are likely F3 and F4, which overlie the dorsolateral prefrontal cortex.Connections
The FEF has extensive connections with other cortical and subcortical regions, including the parietal cortex, superior colliculus, thalamus, and extrastriate visual areas (Schall, 2002; Stanton et al., 2005). These connections are crucial for visual attention, saccadic eye movements, and smooth pursuit.Participation in brain networks
The FEF is a key node in the dorsal attention network responsible for goal-directed attention and eye movement control. This network also includes the intraparietal sulcus and superior parietal lobule (Corbetta & Shulman, 2002).Functions
The FEF is crucial in controlling saccadic eye movements, smooth pursuit, and visual attention. It is involved in the initiation, planning, and execution of eye movements, as well as the allocation of attention to relevant visual stimuli (Schall, 2002). The FEF is vital in cognitive functions, including attention orientation, visual consciousness, access to our conscious experience, perceptual performance, and decision-making (Vernet et al., 2014).Role in clinical disorders
Alterations in FEF function have been implicated in various clinical conditions, including attention deficit hyperactivity disorder (ADHD; Mahone et al., 2011), oculomotor apraxia (Rizzo et al., 1996), and progressive supranuclear palsy (Burrell et al., 2012).Areas 9 and 46: Dorsolateral Prefrontal Cortex (DLPFC)
The dorsolateral prefrontal cortex (DLPFC), which consists of distinct regions, is essential for higher-order cognitive functions, including working memory, executive control, and decision-making (Ahuja & Rodriguez, 2022). Graphic © Big8/Shutterstock.com.Brodmann areas
The DLPFC is primarily located in Brodmann areas 9 and 46 within the lateral aspect of the frontal lobe (Petrides, 2005).Location
The DLPFC is situated in the lateral portion of the frontal lobe, superior and anterior to the premotor and primary motor cortex (M1). The closest sites are likely F3 and F4.Connections
The DLPFC has extensive connections with other cortical and subcortical regions, including the parietal cortex, medial prefrontal cortex, anterior cingulate cortex, orbitofrontal cortex, thalamus, and basal ganglia (Petrides & Pandya, 2002). These connections are critical for cognitive control, working memory, and decision-making.Participation in brain networks
The DLPFC is a key node in the frontoparietal control network, which is responsible for executive control, and the working memory network, which maintains and manipulates information (Cabeza & Nyberg, 2000; Vincent et al., 2008).Functions
The DLPFC plays a crucial role in higher-order cognitive functions such as working memory, executive control, and decision-making. It allocates cognitive resources, goal-directed behavior, task switching, and the flexible adaptation of behavior in response to changing demands (Breedlove & Watson, 2023; Petrides, 2005).Role in clinical disorders
Alterations in DLPFC function have been implicated in various clinical conditions, including schizophrenia (Barch, 2005), attention deficit hyperactivity disorder (ADHD; Cortese et al., 2012), and major depressive disorder (MDD; Drevets et al., 2008).Area 10: Anterior Prefrontal Cortex (aPFC)
The anterior prefrontal cortex (aPFC), also referred to as the frontopolar cortex, is involved in higher-order cognitive processes such as decision-making, planning, and reasoning. Graphic © Big8/Shutterstock.com.Brodmann areas
The aPFC is primarily located in Brodmann area 10, at the most anterior part of the frontal lobe (Ramnani & Owen, 2004).Location
The aPFC is located at the most rostral part of the frontal lobe, anterior to the dorsolateral prefrontal cortex (DLPFC) and orbitofrontal cortex. The closest sites are likely Fp1 and Fp2, which overlie the frontal pole.Connections
The aPFC has extensive connections with other cortical and subcortical regions, including the DLPFC, orbitofrontal cortex, medial prefrontal cortex, posterior parietal cortex, temporal cortex, and thalamus (Burgess et al., 2007). These connections are essential for complex cognitive tasks, multitasking, and mentalizing.Participation in brain networks
The aPFC is a key node in the frontoparietal control network, which is responsible for executive control, as well as the default mode network (DMN), which is involved in self-referential processing and mentalizing (Vincent et al., 2008; Spreng et al., 2009).Functions
The aPFC involves higher-order cognitive processes such as decision-making, planning, reasoning, multitasking, and mentalizing. It is crucial in coordinating and integrating information from various cognitive domains and is responsible for goal-directed behavior and social cognition (Ramnani & Owen, 2004). The aPFC is engaged in various tasks, such as problem-solving, memory recall, future-oriented memory, source and context memory, task-switching, and attention reallocation (Ramnani & Owen, 2004). The aPFC contributes to high-level nociception and pain processing (Peng et al., 2018).Role in clinical disorders
Alterations in aPFC function have been implicated in various clinical conditions, including autism spectrum disorder (ASD; Gilbert et al., 2008), schizophrenia (Perlstein et al., 2001), and major depressive disorder (MDD; Drevets et al., 2008).Areas 11, 12, 13, and 47: Orbitofrontal Cortex (OFC)
The orbitofrontal cortex (OFC) processes reward, emotion, and decision-making and integrates sensory information with emotional valence. Graphics © Big8/Shutterstock.com..jpg)
Brodmann areas
The OFC primarily encompasses Brodmann areas 11, 12, 13, and 47, located in the ventral portion of the frontal lobe (Kringelbach, 2005).Location
The OFC is situated in the ventral part of the frontal lobe, just above the orbits (eye sockets). The anterior prefrontal cortex and the medial prefrontal cortex border it. The closest sites are likely Fp1 and Fp2, which overlie the ventral and rostral portions of the frontal lobe.Connections
The OFC has extensive connections with other cortical and subcortical regions, including the amygdala, insula, cingulate cortex, hippocampus, thalamus, striatum, and sensory cortices (Kringelbach, 2005; Price, 2007). These connections are essential for emotion processing, reward-based decision-making, and social cognition.Participation in brain networks
The OFC is a key node in the salience network, which is responsible for detecting and integrating emotionally and motivationally salient stimuli, and the default mode network (DMN), which is involved in self-referential processing and social cognition (Seeley et al., 2007; Spreng et al., 2009).Functions
The OFC is crucial in processing reward, emotion, and decision-making. It integrates sensory information with emotional valence, evaluates outcomes and actions, and represents social and emotional information (Kringelbach, 2005).Role in clinical disorders
Alterations in OFC function have been implicated in various clinical conditions, including obsessive-compulsive disorder (OCD) (Menzies et al., 2008), major depressive disorder (MDD) (Drevets, 2007), bipolar disorder (BD) (Blumberg et al., 2003), and addiction (Volkow & Fowler, 2000). Depression may be associated with heightened responsiveness and increased connectivity in the lateral orbitofrontal cortex (not linked to rewards), while it is connected to reduced responsiveness and connectivity in the medial orbitofrontal cortex (related to rewards; Rolls, Cheng, & Feng, 2020).Areas 13-16 and 52: Insular Cortex (Insula)
The insular cortex, or insula, is involved in diverse functions, including interoception, emotion processing, pain perception, and cognitive control. Graphics © Big8/Shutterstock.com. Brodmann areas
The insular cortex comprises Brodmann areas 13, 14, 15, 16, and parts of area 52. These include sensorimotor, central-olfactogustatory, socio-emotional, and cognitive anterior-dorsal regions (Kurth et al., 2010).Location
The insular cortex is situated deep within the lateral sulcus, which separates the frontal and parietal lobes from the temporal lobe. The opercula of the frontal, parietal, and temporal lobes cover it.Connections
The insular cortex has extensive connections with various cortical and subcortical regions, including the amygdala, anterior cingulate cortex (ACC), prefrontal cortex, primary and secondary somatosensory cortices, orbitofrontal cortex (OFC), and thalamus (Nieuwenhuys, 2012). These connections contribute to the diverse functions of the insula.Participation in brain networks
The insular cortex is a key node in the salience network, which is responsible for detecting and integrating emotionally and motivationally salient stimuli, and the central autonomic network (CAN), which is involved in autonomic regulation (Seeley et al., 2007; Thayer et al., 2012).Functions
The insular cortex is crucial in interoception, emotion processing, pain perception, and cognitive control. It represents internal bodily states, integrates sensory and emotional information, and modulates cognitive and affective processes (Craig, 2009).Role in clinical disorders
Alterations in insular cortex function have been implicated in various clinical conditions, including anxiety disorders (Paulus & Stein, 2006), major depressive disorder (MDD; Sliz & Hayley, 2012), addiction (Naqvi & Bechara, 2010), and autism spectrum disorder (ASD; Di Martino et al., 2009).Area 17: Primary Visual Cortex (V1)
The primary visual cortex (V1), or the striate cortex, is responsible for processing basic visual information, such as orientation, spatial frequency, and color. Graphic © Big8/Shutterstock.com.Brodmann areas
The primary visual cortex is primarily located in Brodmann area 17, in the occipital lobe (Horton & Adams, 2005).Location
The primary visual cortex is located in the occipital lobe, along the calcarine sulcus, which runs horizontally through the medial part of the lobe. The closest sites are likely O1 and O2, which overlie the occipital lobe.Connections
The primary visual cortex receives input from the lateral geniculate nucleus (LGN) of the thalamus and sends output to the secondary visual cortex (V2) and other extrastriate areas (V3, V4, V5/MT). These connections are essential for the hierarchical processing of visual information (Felleman & Van Essen, 1991).Participation in brain networks
The primary visual cortex is a key node in the visual processing network responsible for processing and interpreting visual information from the retina. This network includes other areas of the occipital lobe and extends to the parietal and temporal cortices (Nassi & Callaway, 2009).Functions
The primary visual cortex processes basic visual information, such as orientation, spatial frequency, and color. It forms the initial stage of the hierarchical processing of visual information and is critical for visual perception (Horton & Adams, 2005).Role in clinical disorders
Alterations in primary visual cortex function have been implicated in various clinical conditions, including amblyopia (lazy eye; Hess et al., 2010), cortical blindness (Celesia, 2005), and visual hallucinations in conditions like Charles Bonnet syndrome (Griffiths, 2000).Areas 18 and 19: Secondary Visual Cortex (V2)
The secondary visual cortex (V2), also known as the prestriate cortex, is involved in the further processing and integration of visual information received from the primary visual cortex (V1). Graphic © Big8/Shutterstock.com.Brodmann areas
The secondary visual cortex is primarily located in Brodmann areas 18 and 19, in the occipital lobe (Tootell et al., 1998).Location
The secondary visual cortex is located in the occipital lobe, surrounding the primary visual cortex along the calcarine sulcus, extending to the lateral parts of the occipital lobe. The closest sites are likely O1 and O2.Connections
The V2 receives input from the primary visual cortex (V1). It sends output to higher-order extrastriate areas (V3, V4, V5/MT) and other cortical regions involved in visual processing, including the parietal and temporal cortices (Felleman & Van Essen, 1991).Participation in brain networks
The secondary visual cortex is a key node in the visual processing network responsible for processing and interpreting visual information from the retina. This network includes other areas of the occipital lobe and parietal and temporal cortices (Nassi & Callaway, 2009).Functions
The secondary visual cortex is involved in further processing and integrating visual information from the primary visual cortex. It is crucial in processing complex visual attributes, such as form, color, and motion (Tootell et al., 1998).Role in clinical disorders
Alterations in secondary visual cortex function have been implicated in various clinical conditions, including visual agnosia, characterized by the inability to recognize objects despite normal visual acuity and intact primary visual cortex function (Milner & Goodale, 2008).Areas 18, 19, 37, 21, and 22: Visual Association Cortex (V3, V4, V5)
The visual association cortex, also known as the higher-order extrastriate cortex, is responsible for the advanced processing of visual information, such as object recognition, face perception, and processing of complex visual scenes. Graphic © Big8/Shutterstock.com.Brodmann areas
The visual association cortex comprises several Brodmann areas, including areas 18, 19, 37, 21, and 22, mainly in the occipital and temporal lobes (Tootell et al., 1998; Kanwisher & Yovel, 2006).Location
The visual association cortex is located primarily in the occipital and temporal lobes, surrounding the primary (V1) and secondary (V2) visual cortices. It includes regions such as the fusiform face area (FFA), the parahippocampal place area (PPA), and the lateral occipital complex (LOC; Epstein & Kanwisher, 1998; Kanwisher & Yovel, 2006; Malach et al., 1995). The closest sites are likely O1, O2, T5, and T6, which overlie the occipital and temporal lobes.Connections
The visual association cortex receives input from the primary (V1) and secondary (V2) visual cortices and has extensive connections with other cortical and subcortical regions, including the parietal lobe, prefrontal cortex, hippocampus, and amygdala (Felleman & Van Essen, 1991; Kravitz et al., 2013).Participation in brain networks
The visual association cortex is a key component of the ventral visual processing stream, also known as the "what" pathway, responsible for object recognition and processing of complex visual scenes (Kravitz et al., 2011).Functions
The visual association cortex is involved in advanced visual processing, including object recognition, face perception, processing of complex visual scenes, and integration of visual information with other sensory modalities (Kanwisher & Yovel, 2006; Tootell et al., 1998).Role in clinical disorders
Alterations in visual association cortex function have been implicated in various clinical conditions, including prosopagnosia (face blindness; Duchaine & Nakayama, 2006), visual agnosia (Milner & Goodale, 2008), and higher-order visual processing deficits in conditions such as autism spectrum disorder (ASD; Simmons et al., 2009).Areas 20 and 37: Inferior Temporal Gyrus (ITG)
The inferior temporal gyrus (ITG) is a part of the temporal lobe involved in high-level visual processing and object recognition. Graphic © Big8/Shutterstock.com.Brodmann areas
The inferior temporal gyrus primarily includes Brodmann areas 20 and 37 (Amunts et al., 2000).Location
The inferior temporal gyrus is located in the ventral part of the temporal lobe, below the middle temporal gyrus and superior temporal sulcus, and above the fusiform gyrus. The closest sites are likely T5 (or TP7) and T6 (or TP8), which overlie the temporal lobes.Connections
The ITG has extensive connections with other cortical and subcortical regions, including the primary and secondary visual cortices, fusiform gyrus, parahippocampal gyrus, hippocampus, amygdala, and prefrontal cortex (Kravitz et al., 2013; Felleman & Van Essen, 1991).Participation in brain networks
The ITG is a key component of the ventral visual processing stream, also known as the "what" pathway, responsible for object recognition and processing of complex visual scenes (Kravitz et al., 2011).Functions
The ITG is involved in high-level visual processing, object recognition, semantic processing, and the integration of visual information with other sensory modalities (Kanwisher & Yovel, 2006).Role in clinical disorders
Alterations in ITG function have been implicated in various clinical conditions, including visual agnosia, prosopagnosia (face blindness), and higher-order visual processing deficits in conditions such as autism spectrum disorder (ASD; Duchaine & Nakayama, 2006; Simmons et al., 2009).Areas 21 and 39: Middle Temporal Gyrus (MTG)
The middle temporal gyrus (MTG) is a part of the temporal lobe involved in various functions, such as semantic processing, language, and high-level visual processing. Graphics © Big8/Shutterstock.com. Brodmann areas
The MTG primarily includes Brodmann areas 21 and 39 (Amunts et al., 2000).Location
The MTG is located in the lateral part of the temporal lobe, between the superior temporal gyrus (above) and the inferior temporal gyrus (below), and adjacent to the superior temporal sulcus.Connections
The MTG has extensive connections with other cortical and subcortical regions, including the primary and secondary visual cortices, the angular gyrus, the fusiform gyrus, the parahippocampal gyrus, the hippocampus, the amygdala, and the prefrontal cortex (Kravitz et al., 2013; Felleman & Van Essen, 1991).Participation in brain networks
The MTG participates in various brain networks, including the ventral visual processing stream ("what" pathway) for object recognition and processing of complex visual scenes (Kravitz et al., 2011), and the language network for semantic processing and word retrieval (Binder et al., 2009).Functions
The MTG is involved in various functions, such as semantic processing, language comprehension, word retrieval, and high-level visual processing, including object and face recognition (Binder et al., 2009; Kanwisher & Yovel, 2006).Role in clinical disorders
Alterations in MTG function have been implicated in various clinical conditions, including semantic dementia (Hodges et al., 1992), language impairments in aphasia (Dronkers et al., 2004), and higher-order visual processing deficits in conditions such as autism spectrum disorder (ASD; Simmons et al., 2009).Areas 22, 39, and 40: Superior Temporal Gyrus (STG)
The superior temporal gyrus (STG), including Wernicke's area, is a part of the temporal lobe involved in various functions such as language comprehension, auditory processing, and social cognition. Graphic © Big8/Shutterstock.com.Brodmann areas
Wernicke's area primarily includes Brodmann area 22 and, to some extent, areas 39 and 40 (Amunts et al., 2000).Location
Wernicke's area is located in the posterior part of the superior temporal gyrus, usually in the left hemisphere, near the lateral sulcus. The STG runs laterally along the temporal lobe, above the middle temporal gyrus. The closest site is likely T5 (or TP7) for the left hemisphere, where Wernicke's area is typically located.Connections
Wernicke's area has extensive connections with other language-related regions, including Broca's area (via the arcuate fasciculus), the angular gyrus, and other parts of the superior temporal gyrus (Friederici, 2009). The STG also connects with the primary and secondary auditory cortices, social cognition, and memory regions.Participation in brain networks
Wernicke's area participates in the language network, playing a crucial role in language comprehension and semantic processing (Binder et al., 2009). The STG is also involved in the auditory processing network and the social cognition network.Functions
Wernicke's area involves language comprehension, semantic processing, and integrating auditory information into meaningful speech (Price, 2012). The STG also plays a role in auditory processing, social cognition, and memory.Role in clinical disorders
Alterations in the function of Wernicke's area and the STG have been implicated in various clinical conditions, such as Wernicke's aphasia, characterized by impaired language comprehension and fluent but nonsensical speech (Dronkers et al., 2004). The STG has also been implicated in auditory processing deficits and social cognition impairments in conditions such as autism spectrum disorder (ASD; Boddaert et al., 2004).Area 23: Ventral Posterior Cingulate Cortex (vPCC)
The ventral posterior cingulate cortex (vPCC) is a region within the posterior cingulate cortex (PCC), a part of the limbic system involved in various functions such as memory, emotion, and self-referential processing. Graphic © Big8/Shutterstock.com. Brodmann areas
The vPCC primarily includes Brodmann area 23 (Vogt et al., 2006).Location
The vPCC is located in the medial part of the brain, in the posterior cingulate cortex, and ventral to the dorsal posterior cingulate cortex (dPCC). It is positioned between the precuneus and the corpus callosum. The closest sites are likely Pz and CPz, located over the midline parietal and central regions, respectively.Connections
The vPCC connects with various brain regions, including the medial prefrontal cortex (mPFC), hippocampus, parahippocampal gyrus, and lateral parietal regions (Leech & Sharp, 2014; Utevsky et al., 2014).Participation in brain networks
The vPCC is a key component of the default mode network (DMN), which is active during rest and involved in self-referential thinking, autobiographical memory, and social cognition (Raichle et al., 2001; Buckner et al., 2008).Functions
The vPCC is involved in various functions, such as self-referential thinking, autobiographical memory, social cognition, and emotional processing (Leech & Sharp, 2014; Utevsky et al., 2014).Role in clinical disorders
Alterations in vPCC function have been implicated in various clinical conditions, including Alzheimer's disease (Buckner et al., 2005), major depressive disorder (Sheline et al., 2010), and autism spectrum disorder (ASD; Padmanabhan et al., 2017).Areas 24 and 25: Ventral Anterior Cingulate Cortex (vACC)
The ventral anterior cingulate cortex (vACC) is a region within the anterior cingulate cortex (ACC), which is part of the limbic system and involved in various functions, such as emotion processing, reward-based learning, and decision-making. Graphic © Big8/Shutterstock.com. Brodmann areas
The vACC primarily includes Brodmann areas 24 and 25 (Vogt, 2005).Location
The vACC is located in the medial part of the brain, in the anterior cingulate cortex, ventral to the dorsal anterior cingulate cortex (dACC). It is positioned anterior to the genu of the corpus callosum. The closest sites are likely FCz and Cz, located over the midline frontal and central regions, respectively.Connections
The vACC has connections with various brain regions, including the amygdala, hippocampus, medial prefrontal cortex (mPFC), orbitofrontal cortex (OFC), and nucleus accumbens (Bush et al., 2000; Etkin et al., 2011).Participation in brain networks
The vACC is a key component of the salience network, which detects and integrates salient emotional and sensory stimuli and modulates attention and cognitive control (Menon, 2011; Seeley et al., 2007).Functions
The vACC is implicated in various cognitive and emotional functions, including error detection, conflict monitoring, emotion regulation, empathy, and social cognition (Bush et al., 2000; Etkin et al., 2011).Role in clinical disorders
Abnormalities in the vACC have been implicated in several psychiatric and neurological disorders, such as depression, anxiety, schizophrenia, bipolar disorder, attention deficit hyperactivity disorder (ADHD), and autism spectrum disorders (Drevets et al., 2008; Etkin et al., 2010).Areas 25 and 24b: Subgenual Ventromedial Prefrontal Cortex (vmPFC)
The subgenual region of the ventromedial prefrontal cortex (vmPFC) is an important brain region involved in various cognitive and emotional processes. Graphic © Big8/Shutterstock.com. Brodmann areas
The subgenual region of the vmPFC primarily consists of Brodmann areas 25 and 24b (Ongür et al., 2003).Location
The subgenual region of the vmPFC is located in the medial prefrontal cortex, ventral to the genu of the corpus callosum, and adjacent to the anterior cingulate cortex (Mayberg, 2003). Its nearby EEG electrode positions include Fp1, Fp2, Fz, and AFz, located along the scalp's midline (Jasper, 1958).Connections
The subgenual region of the vmPFC has extensive connections with other brain regions, including the amygdala, hippocampus, hypothalamus, nucleus accumbens, thalamus, and other prefrontal areas (Ongür et al., 2003; Price & Drevets, 2010).Participation in brain networks
The subgenual vmPFC is a key component of the default mode network (DMN) and the affective network, involved in self-referential processing, emotion regulation, and decision-making (Buckner et al., 2008; Rudebeck et al., 2014).Functions
The subgenual vmPFC is implicated in various cognitive and emotional functions, including value-based decision-making, emotion regulation, self-referential processing, and social cognition (Rudebeck et al., 2014; Roy et al., 2012).Role in clinical disorders
Abnormalities in the subgenual vmPFC have been implicated in several psychiatric disorders, such as major depressive disorder, bipolar disorder, anxiety disorders, and post-traumatic stress disorder (Mayberg, 2003; Price & Drevets, 2010).Areas 29 and 30: Ectosplenial Retrosplenial Cerebral Cortex
The ectosplenial region is not a widely recognized or well-established region within the human retrosplenial cortex. However, the retrosplenial cortex is a crucial brain area involved in various cognitive processes, particularly spatial memory, and navigation. Graphic © Big8/Shutterstock.com. Brodmann areas
The retrosplenial cortex mainly comprises Brodmann areas 29 and 30, in the posterior cingulate cortex (Vogt et al., 2006).Location
The retrosplenial cortex is situated in the medial parietal lobe, posterior to the splenium of the corpus callosum, and adjacent to the posterior cingulate cortex (Vann et al., 2009). Its nearby EEG electrode positions include Pz, CPz, and Oz, located along the midline of the scalp (Jasper, 1958).Connections
The retrosplenial cortex has extensive connections with other brain regions, including the hippocampus, parahippocampal cortex, thalamus, anterior cingulate cortex, and other parietal and frontal areas (Vann et al., 2009).Participation in brain networks
The retrosplenial cortex is a key component of the default mode network (DMN) and is involved in spatial memory, episodic memory, and self-referential processing (Buckner et al., 2008).Functions
The retrosplenial cortex is implicated in various cognitive functions, including spatial memory, navigation, episodic memory, and scene construction (Epstein, 2008; Vann et al., 2009).Role in clinical disorders
Abnormalities in the retrosplenial cortex have been implicated in several neurological and psychiatric disorders, such as Alzheimer's disease, amnesia, and schizophrenia, which often involve impairments in spatial memory and navigation (Maguire, 2001; Mendez & Cherrier, 2003).Areas 29 and 30: Retrosplenial Cingulate Cortex
The retrosplenial cingulate cortex is an important brain region involved in various cognitive processes, particularly related to spatial memory and navigation. Graphic © Big8/Shutterstock.com. Brodmann areas
The retrosplenial cortex mainly comprises Brodmann areas 29 and 30, which are located in the posterior cingulate cortex (Vogt et al., 2006).Location
The retrosplenial cortex is situated in the medial parietal lobe, posterior to the splenium of the corpus callosum, and adjacent to the posterior cingulate cortex (Vann et al., 2009). Its nearby EEG electrode positions include Pz, CPz, and Oz, located along the midline of the scalp (Jasper, 1958).Connections
The retrosplenial cortex has extensive connections with other brain regions, including the hippocampus, parahippocampal cortex, thalamus, anterior cingulate cortex, and other parietal and frontal areas (Vann et al., 2009).Participation in brain networks
The retrosplenial cortex is a key component of the default mode network (DMN) and is involved in spatial memory, episodic memory, and self-referential processing (Buckner et al., 2008).Functions
The retrosplenial cortex is implicated in various cognitive functions, including spatial memory, navigation, episodic memory, and scene construction (Vann et al., 2009; Epstein, 2008).Role in clinical disorders
Abnormalities in the retrosplenial cortex have been implicated in several neurological and psychiatric disorders, such as Alzheimer's disease, amnesia, and schizophrenia, which often involve impairments in spatial memory and navigation (Maguire, 2001; Mendez & Cherrier, 2003).Areas 23, 24, and 31: Dorsal Posterior Cingulate Cortex (dPCC)
The dorsal posterior cingulate cortex (dPCC) is an important brain region involved in various cognitive processes, particularly related to attention and memory. Graphic © Big8/Shutterstock.com. Brodmann areas
The dPCC is primarily composed of Brodmann areas 23 and 31. These areas are associated with the cingulate cortex and form part of the limbic system, which plays a crucial role in emotion formation, processing, learning, and memory (Vogt, Finch, & Olson, 1992).Location
The dPCC is located in the medial aspect of the brain, towards the back. It's located directly above the corpus callosum, a nerve fiber bundle connecting the left and right cerebral hemispheres.Connections
The dPCC has numerous connections with other areas of the brain. It connects with other regions of the cingulate cortex, the medial prefrontal cortex, the parahippocampal gyrus, and the precuneus. It also connects with the thalamus and various parts of the temporal and parietal lobes. These connections make the dPCC a central hub for information processing and transfer (Margulies et al., 2009).Participation in brain networks
The dPCC is part of several crucial brain networks. It is an integral part of the default mode network (DMN), which is most active when the brain is at rest and not focused on the outside world. The dPCC also interacts with the salience network, which is crucial for determining the sensory or emotional inputs most relevant to our goals and current situation (Leech & Sharp, 2014).Functions
The functions of the dPCC are diverse and complex due to its involvement in various brain networks and its wide-ranging connections. These functions include self-referential thought, episodic memory retrieval, and consciousness. It also plays a role in internally directed thought, such as daydreaming, future planning, and moral reasoning (Andrews-Hanna et al., 2010).Role in clinical disorders
Abnormalities or dysfunction in the dPCC have been linked to several clinical disorders. These include Alzheimer's disease, where decreased activity in the dPCC has been associated with the early stages of the disease (Buckner, R. L., et al., 2005). The dPCC has also been implicated in various psychiatric disorders, such as depression, anxiety, and schizophrenia, where altered connectivity within and between networks involving the dPCC is often seen (Greicius et al., 2007).Areas 24, 25, 32, and 33: Anterior Cingulate Cortex (ACC)
The anterior cingulate cortex (ACC) is a crucial brain region involved in various cognitive, emotional, and regulatory processes. Graphic © Big8/Shutterstock.com.Brodmann areas
The ACC is divided into several subregions, including the dorsal ACC (dACC; Brodmann areas 24 and 32) and the ventral ACC (vACC; Brodmann areas 25 and 33; Vogt, 2009).Location
The ACC is located in the medial aspect of the cerebral cortex, surrounding the corpus callosum, with the dACC situated dorsally and the vACC situated ventrally (Vogt, 2009).Connections
The ACC has extensive connections with other brain regions, including the prefrontal cortex, parietal cortex, amygdala, hippocampus, thalamus, and other limbic areas (Devinsky et al., 1995).Participation in brain networks
The ACC is a key component of several brain networks, including the default mode network (DMN), the salience network, and the executive control network, which are involved in cognitive, emotional, and behavioral processing (Bressler & Menon, 2010).Functions
The ACC is implicated in various cognitive functions, including attention, error detection, conflict monitoring, emotion regulation, and decision-making (Bush et al., 2000).Role in clinical disorders
Abnormalities in the ACC have been implicated in several neurological and psychiatric disorders, such as Alzheimer's disease, depression, anxiety, and schizophrenia, which often involve impairments in cognitive, emotional, and behavioral processing (Vogt, 2005).Areas 24, 32, and 33: Cingulate Cortex
The cingulate cortex is an important brain region involved in various cognitive, emotional, and behavioral processes. Graphic © Big8/Shutterstock.com.
Brodmann areas
The cingulate cortex is divided into several subregions, including the anterior cingulate cortex (ACC; Brodmann areas 24, 32, and 33) and the posterior cingulate cortex (PCC; Brodmann areas 23, 29, and 30; Vogt, 2009).Location
The cingulate cortex is located in the medial aspect of the cerebral cortex, surrounding the corpus callosum, with the ACC situated anteriorly and the PCC situated posteriorly (Vogt, 2009). EEG electrode positions near the cingulate cortex include Fz, FCz, and Cz, located along the scalp's midline (Jasper, 1958).Connections
The cingulate cortex has extensive connections with other brain regions, including the prefrontal cortex, parietal cortex, amygdala, hippocampus, thalamus, and other limbic areas (Devinsky et al., 1995).Participation in brain networks
The cingulate cortex is a key component of several brain networks, including the default mode network (DMN), the salience network, and the executive control network, which are involved in cognitive, emotional, and behavioral processing (Bressler & Menon, 2010).Functions
The cingulate cortex is implicated in various cognitive functions, including attention, error detection, conflict monitoring, emotion regulation, and decision-making (Bush et al., 2000).Role in clinical disorders
Abnormalities in the cingulate cortex have been implicated in several neurological and psychiatric disorders, such as Alzheimer's disease, depression, anxiety, and schizophrenia, which often involve impairments in cognitive, emotional, and behavioral processing (Vogt, 2005).Area 27: Pyriform (Piriform) Cortex
The pyriform cortex, also known as the primary olfactory cortex, is a crucial brain region that processes olfactory information. Graphic © Big8/Shutterstock.com. Brodmann areas
The pyriform cortex is not typically associated with specific Brodmann areas. It is part of the allocortex, which has a simpler organization than the isocortex, where Brodmann areas are usually defined (Shepherd, 2007).Location
The pyriform cortex is in the medial temporal lobe, anterior to the perirhinal cortex and lateral to the amygdala (Neville & Haberly, 2004).Connections
The pyriform cortex has extensive connections with other brain regions, including the olfactory bulb, amygdala, thalamus, orbitofrontal cortex, and hippocampus, which are involved in processing and integrating olfactory information (Gottfried, 2010).Participation in brain networks
The pyriform cortex is a key component of the olfactory network, which processes and integrates olfactory information from the environment and plays a role in memory, emotion, and decision-making (Gottfried, 2010).Functions
The pyriform cortex primarily processes olfactory information, including odor discrimination, odor memory, and odor-guided behavior (Neville & Haberly, 2004).Role in clinical disorders
Abnormalities in the piriform cortex have been implicated in several neurological and psychiatric disorders, such as Alzheimer's disease, Parkinson's disease, and schizophrenia, which often involve impairments in olfactory function (Doty, 2008).Area 28: Ventral Entorhinal Cortex (vEC)
The ventral entorhinal cortex (vEC) is an important brain region for various cognitive processes, particularly memory and spatial navigation. Graphic © Big8/Shutterstock.com. Brodmann areas
The entorhinal cortex is not typically associated with specific Brodmann areas, as it is part of the allocortex, which has a simpler organization than the isocortex, where Brodmann areas are usually defined (Witter et al., 2000).Location
The ventral entorhinal cortex is located in the medial temporal lobe, anterior to the hippocampus and posterior to the perirhinal cortex (Witter et al., 2000).Connections
The ventral entorhinal cortex has extensive connections with other brain regions, including the hippocampus, perirhinal cortex, parahippocampal cortex, and prefrontal cortex, which are involved in memory and spatial navigation (van Strien et al., 2009; Witter et al., 2000).Participation in brain networks
The ventral entorhinal cortex is a key component of the medial temporal lobe memory system, crucial for episodic memory and spatial navigation (Eichenbaum et al., 2007).Functions
The ventral entorhinal cortex is implicated in various cognitive functions, including episodic memory and spatial navigation (Eichenbaum et al., 2007; Hafting et al., 2005).Role in clinical disorders
Abnormalities in the ventral entorhinal cortex have been implicated in several neurological and psychiatric disorders, such as Alzheimer's disease, temporal lobe epilepsy, and schizophrenia, which often involve impairments in memory and spatial navigation (Braak & Braak, 1991; Du et al., 2017).Areas 28 and 34: Dorsal Entorhinal Cortex (dEC)
The dorsal entorhinal cortex (dEC) is an important brain region in spatial memory and navigation. Graphic © Big8/Shutterstock.com. Brodmann areas
Brodmann areas do not easily define the entorhinal cortex (EC), as it is an evolutionarily conserved structure that does not map neatly onto the cytoarchitectonic divisions. However, it is often associated with Brodmann areas 28 and 34 (Van Strien et al., 2009).Location
The dEC is located in the medial temporal lobe, situated dorsal to the ventral entorhinal cortex (vEC) (Van Strien et al., 2009).Connections
The dEC has extensive connections with other brain regions, including the hippocampus, perirhinal cortex, parahippocampal cortex, and other medial temporal lobe structures (Witter et al., 2000).Participation in brain networks
The dEC is involved in the medial temporal lobe memory system, which plays a crucial role in spatial memory and navigation (Eichenbaum, 2000).Functions
The dEC is implicated in various cognitive functions, including spatial memory, navigation, and contextual processing (Hafting et al., 2005).Role in clinical disorders
Abnormalities in the dEC have been implicated in several neurological disorders, such as Alzheimer's, which involves memory and navigation impairments (Khan et al., 2014).Areas 29 and 30: Ectosplenial Retrosplenial Cerebral Cortex
The ectosplenial region is not a widely recognized or well-established region within the human retrosplenial cortex. However, the retrosplenial cortex is a crucial brain area involved in various cognitive processes, particularly spatial memory and navigation. Graphic © Big8/Shutterstock.com. Brodmann areas
The retrosplenial cortex mainly comprises Brodmann areas 29 and 30, located in the posterior cingulate cortex (Vogt et al., 2006).Location
The retrosplenial cortex is situated in the medial parietal lobe, posterior to the splenium of the corpus callosum, and adjacent to the posterior cingulate cortex (Vann et al., 2009). Its nearby EEG electrode positions include Pz, CPz, and Oz, located along the midline of the scalp (Jasper, 1958).Connections
The retrosplenial cortex has extensive connections with other brain regions, including the hippocampus, parahippocampal cortex, thalamus, anterior cingulate cortex, and other parietal and frontal areas (Vann et al., 2009).Participation in brain networks
The retrosplenial cortex is a key component of the default mode network (DMN) and is involved in spatial memory, episodic memory, and self-referential processing (Buckner et al., 2008).Functions
The retrosplenial cortex is implicated in various cognitive functions, including spatial memory, navigation, episodic memory, and scene construction (Epstein, 2008; Vann et al., 2009).Role in clinical disorders
Abnormalities in the retrosplenial cortex have been implicated in several neurological and psychiatric disorders, such as Alzheimer's disease, amnesia, and schizophrenia, which often involve impairments in spatial memory and navigation (Maguire, 2001; Mendez & Cherrier, 2003).Areas 35 and 36: Perirhinal Cortex (PRC)
The perirhinal cortex (PRC) is a significant brain region involved in various cognitive processes, such as object recognition and memory. Graphic © Big8/Shutterstock.com.Brodmann areas
The PRC is associated with Brodmann areas 35 and 36, located within the medial temporal lobe (Van Hoesen & Pandya, 1975).Location
The PRC is located in the medial temporal lobe, adjacent to the entorhinal and parahippocampal cortex (Van Hoesen & Pandya, 1975).Connections
The PRC has extensive connections with other brain regions, including the entorhinal cortex, hippocampus, parahippocampal cortex, amygdala, and other medial temporal lobe structures (Suzuki & Amaral, 1994).Participation in brain networks
The PRC is a crucial component of the medial temporal lobe memory system, playing an essential role in object recognition, associative memory, and familiarity-based recognition (Eichenbaum et al., 2007).Functions
The PRC is implicated in various cognitive functions, including object recognition, associative memory, and familiarity-based recognition (Eichenbaum et al., 2007).Role in clinical disorders
Abnormalities in the PRC have been implicated in several neurological disorders, such as Alzheimer's disease and other memory-related disorders (Khan et al., 2014).Areas 37 and 19: Fusiform Gyrus
The fusiform gyrus is a key brain region involved in various cognitive processes, such as face and object recognition. Graphic © Big8/Shutterstock.com.Brodmann areas
The fusiform gyrus is associated with Brodmann areas 37 and 19, located on the ventral surface of the temporal and occipital lobes (Grill-Spector et al., 2001).Location
The fusiform gyrus is located on the ventral surface of the temporal and occipital lobes, medial to the inferior temporal gyrus, and lateral to the parahippocampal gyrus (Grill-Spector et al., 2001).Connections
The fusiform gyrus has extensive connections with other brain regions, including the inferior temporal cortex, occipital cortex, parietal cortex, amygdala, and other medial temporal lobe structures (Catani et al., 2003).Participation in brain networks
The fusiform gyrus is involved in the ventral visual processing stream, playing a crucial role in object and face recognition and other high-level visual processes (Grill-Spector et al., 2001).Functions
The fusiform gyrus is implicated in various cognitive functions, including object recognition, face recognition, and high-level visual processing (Grill-Spector et al., 2001).Role in clinical disorders
Abnormalities in the fusiform gyrus have been implicated in several neurological disorders, such as prosopagnosia, autism spectrum disorders, and Alzheimer's disease (Avidan & Behrmann, 2009).Area 38: Temporopolar Area (Temporal Pole)
The temporopolar area, also known as the temporal pole, involves various cognitive and emotional processes. Graphic © Big8/Shutterstock.com.Brodmann areas
The temporopolar area is associated with Brodmann area 38, located at the most anterior part of the temporal lobe (Öngür et al., 2003).Location
The temporopolar area is located at the most anterior part of the temporal lobe, anterior to the superior, middle, and inferior temporal gyri (Öngür et al., 2003).Connections
The temporopolar area has extensive connections with other brain regions, including the amygdala, hippocampus, orbitofrontal cortex, insula, and other temporal lobe structures (Olson et al., 2007).Participation in brain networks
The temporopolar area is involved in various brain networks, including the default mode and salience networks, playing crucial roles in social cognition, emotional processing, and semantic memory (Roy et al., 2009).Functions
The temporopolar area is implicated in various cognitive functions, including social cognition, emotional processing, and semantic memory (Roy et al., 2009).Role in clinical disorders
Abnormalities in the temporopolar area have been implicated in several neurological disorders, such as frontotemporal dementia, Alzheimer's disease, and other memory-related disorders (Seeley et al., 2009).Area 39: Angular Gyrus
The angular gyrus is involved in various cognitive processes, such as language, attention, and spatial cognition. Graphic © Big8/Shutterstock.com.Brodmann areas
The angular gyrus is associated with Brodmann area 39 in the parietal lobe (Caspers et al., 2006).Location
The angular gyrus is located in the parietal lobe, at the junction of the superior temporal and occipital lobes, and is bordered by the supramarginal gyrus and the occipital cortex (Caspers et al., 2006). The angular gyrus is near the P3 and P4 electrode sites of the International 10-20 system (Jasper, 1958).Connections
The angular gyrus has extensive connections with other brain regions, including the prefrontal cortex, posterior cingulate cortex, superior temporal sulcus, and other regions within the parietal lobe (Seghier, 2013).Participation in brain networks
The angular gyrus plays crucial roles in attention, memory, and language processing in several brain networks, such as the default mode and frontoparietal control networks (Seghier, 2013).Functions
The angular gyrus is implicated in various cognitive functions, including language processing, attention, spatial cognition, and mathematical processing (Seghier, 2013).Role in clinical disorders
Abnormalities in the angular gyrus have been implicated in several neurological disorders, such as dyslexia, aphasia, and Gerstmann syndrome, which involve impairments in language, calculation, and other cognitive processes (Hoeft et al., 2007).Area 40: Supramarginal Gyrus
The supramarginal gyrus is a brain region involved in various cognitive processes, such as language, attention, and sensorimotor integration. Graphic © Big8/Shutterstock.com.Brodmann areas
The supramarginal gyrus is associated with Brodmann area 40, located in the parietal lobe (Caspers et al., 2006).Location
The supramarginal gyrus is located in the parietal lobe at the posterior end of the Sylvian fissure and is bordered by the angular gyrus and the postcentral gyrus (Caspers et al., 2006). It is situated near the P3 and P4 electrode sites of the International 10-20 system (Jasper, 1958).Connections
The supramarginal gyrus is involved in several brain networks, such as the frontoparietal control and dorsal attention networks, playing crucial roles in attention, language processing, and sensorimotor integration (Caspers et al., 2011).Participation in brain networks
The supramarginal gyrus is involved in several brain networks, such as the frontoparietal control network and the dorsal attention network, playing crucial roles in attention, language processing, and sensorimotor integration (Caspers et al., 2011).Functions
The supramarginal gyrus is implicated in various cognitive functions, including language processing, attention, sensorimotor integration, and working memory (Caspers et al., 2011).Role in clinical disorders
Abnormalities in the supramarginal gyrus have been implicated in several neurological disorders, such as dyslexia, apraxia, and other cognitive impairments involving language and sensorimotor processing (Hoeft et al., 2007).Areas 41 and 42: Auditory Cortex
The auditory cortex is a brain region involved in processing auditory information. Graphic © Big8/Shutterstock.com.Brodmann areas
The auditory cortex comprises several Brodmann areas, including the primary auditory cortex (Brodmann areas 41 and 42) and the surrounding secondary auditory cortex (Brodmann area 22; Morosan et al., 2001).Location
The auditory cortex is located in the superior temporal gyrus within the Sylvian fissure, extending into the lateral sulcus in the temporal lobe (Morosan et al., 2001).Connections
The auditory cortex has extensive connections with other brain regions, including the thalamus (specifically, the medial geniculate nucleus), inferior colliculus, and other cortical regions involved in language, attention, and multisensory integration (Bizley & Cohen, 2013).Participation in brain networks
The auditory cortex participates in several brain networks, such as the auditory processing, language, and attention networks, playing crucial roles in sound processing, speech perception, and auditory attention (Griffiths & Warren, 2002).Functions
The auditory cortex involves various functions, including sound processing, speech perception, scene analysis, and attention (Griffiths & Warren, 2002).Role in clinical disorders
Abnormalities in the auditory cortex have been implicated in several neurological disorders, such as tinnitus, auditory processing disorders, and language-related impairments like dyslexia (Sedley et al., 2015).Area 43: Primary Gustatory Cortex (PGC)
The primary gustatory cortex (PGC) is a brain region that processes taste information. Graphic © Big8/Shutterstock.com.Brodmann areas
The PGC is associated with Brodmann area 43, also known as the opercular part of the inferior frontal gyrus and part of the insular cortex (Brodmann area 13; Ogawa, 2012; Small et al., 1999).Location
The PGC is situated in the insular cortex, specifically in the anterior insula, and extends into the adjacent opercular part of the inferior frontal gyrus (Small et al., 1999).Connections
The PGC connects with various brain regions, including the thalamus (specifically, the ventroposteromedial nucleus), orbitofrontal cortex, amygdala, and other cortical regions involved in multisensory integration, emotion, and memory (Rolls, 2006).Participation in brain networks
The PGC is part of the gustatory processing network, which involves taste perception and associated emotional and cognitive processes (Rolls, 2006).Functions
The PGC processes taste information, including taste perception, taste discrimination, and integration with other sensory modalities (Small et al., 1999).Role in clinical disorders
Abnormalities in the PGC have been implicated in several neurological disorders, such as taste-related disorders (ageusia) and eating disorders (anorexia nervosa; Frank et al., 2016).Area 44: Pars Opercularis (inferior temporal gyrus and part of Broca's area)
The pars opercularis is a brain region involved in language processing and motor control. Graphic © Big8/Shutterstock.com.Brodmann areas
The pars opercularis is part of Brodmann area 44, also known as the opercular part of the inferior frontal gyrus (Amunts et al., 1999).Location
The pars opercularis is situated in the inferior frontal gyrus, posterior to the pars triangularis, and anterior to the precentral gyrus in the frontal lobe (Amunts et al., 1999). It is located near the F7 and F8 electrode sites of the International 10-20 system (Jasper, 1958).Connections
The pars opercularis connects with various brain regions, including the posterior superior temporal gyrus (Wernicke's area), precentral gyrus, supplementary motor area, and other cortical regions involved in language processing and motor control (Friederici, 2011).Participation in brain networks
The pars opercularis is part of the language and motor networks, playing crucial roles in speech production, syntactic processing, and motor control (Friederici, 2011).Functions
The pars opercularis is involved in various functions, including speech production, syntactic processing, and motor control (Friederici, 2011).Role in clinical disorders
Abnormalities in the pars opercularis have been implicated in several neurological disorders, such as developmental language disorders, stuttering, and apraxia of speech (Neef et al., 2018; Watkins et al., 2002).Area 45: Pars Triangularis (inferior temporal gyrus and part of Broca's area)
The pars triangularis is a brain region involved in language processing and executive functions. Graphic © Big8/Shutterstock.com.
Brodmann areas
The pars triangularis is part of Brodmann area 45, also known as the triangular part of the inferior frontal gyrus (Amunts et al., 1999).Location
The pars triangularis is situated in the inferior frontal gyrus, anterior to the pars opercularis, and posterior to the pars orbitalis in the frontal lobe (Amunts et al., 1999). It is located near the F7 and F8 electrode sites of the International 10-20 system (Jasper, 1958).Connections
The pars triangularis connects with various brain regions, including the posterior superior temporal gyrus (Wernicke's area), dorsolateral prefrontal cortex, anterior cingulate cortex, and other cortical regions involved in language processing and executive functions (Friederici, 2011).Participation in brain networks
The pars triangularis is part of the language and executive control networks, playing crucial roles in semantic processing, working memory, and cognitive control (Friederici, 2011).Functions
The pars triangularis is involved in various functions, including semantic processing, working memory, and cognitive control (Friederici, 2011).Role in clinical disorders
Abnormalities in the pars triangularis have been implicated in several neurological disorders, such as developmental language disorders, aphasia, and ADHD (Booth et al., 2005; Watkins et al., 2002).Areas 9, 46, 8, and 10: Dorsolateral Prefrontal Cortex (DLPFC)
The dorsolateral prefrontal cortex (DLPFC) is a critical brain region involved in various cognitive and executive functions. Graphic © Big8/Shutterstock.com.
Brodmann areas
The DLPFC mainly includes Brodmann areas 9 and 46 and parts of areas 8 and 10 (Rajkowska & Goldman-Rakic, 1995).Location
The DLPFC is situated in the lateral and superior part of the frontal lobe, encompassing the middle and superior frontal gyri (Rajkowska & Goldman-Rakic, 1995). The DLPFC is located near the F3 and F4 electrode sites of the International 10-20 system (Jasper, 1958).Connections
The DLPFC connects with various brain regions, including the parietal cortex, anterior cingulate cortex, thalamus, and striatum, forming key nodes within the fronto-parietal and cingulo-opercular networks (Fuster, 2001; Dosenbach et al., 2007).Participation in brain networks
The DLPFC is part of the central executive network, playing crucial roles in cognitive control, working memory, decision-making, and goal-directed behavior (Fuster, 2001; Niendam et al., 2012).Functions
The DLPFC is involved in various functions, including cognitive control, working memory, decision-making, and goal-directed behavior (Fuster, 2001; Niendam et al., 2012).Role in clinical disorders
Abnormalities in the DLPFC have been implicated in several neurological disorders, such as schizophrenia, depression, and ADHD (Broyd et al., 2009; Cao et al., 2021; Liston et al., 2011).Area 47: Pars Orbitalis (part of the inferior frontal gyrus)
The pars orbitalis is a brain region involved in various cognitive and emotional processes. Graphic © Big8/Shutterstock.com.
Brodmann areas
The pars orbitalis is part of Brodmann area 47, located in the orbital part of the inferior frontal gyrus (Amunts et al., 1999).Location
The pars orbitalis is situated in the inferior frontal gyrus, anterior to the pars triangularis, and posterior to the lateral orbital gyrus in the frontal lobe (Amunts et al., 1999). It is located near the Fp1 and Fp2 electrode sites.Connections
The pars orbitalis connects with various brain regions, including the amygdala, insula, anterior cingulate cortex, and other cortical regions involved in emotional processing, decision-making, and social cognition (Barbas, 2007; Ongür & Price, 2000).Participation in brain networks
The pars orbitalis is part of the salience network and other networks associated with emotional processing, decision-making, and social cognition (Seeley et al., 2007).Functions
The pars orbitalis involves various functions, including emotional processing, decision-making, and social cognition (Barbas, 2007; Ongür & Price, 2000).Role in clinical disorders
Abnormalities in the pars orbitalis have been implicated in several neurological disorders, such as mood disorders, anxiety disorders, and autism spectrum disorders (Phillips et al., 2003; Di Martino et al., 2009).Area 48: Retrosubicular Area (small medial temporal lobe area)
The retrosubicular area, also called the presubiculum, is a part of the hippocampal formation involved in various cognitive processes, particularly spatial navigation and memory. Graphic © Big8/Shutterstock.com.
Brodmann areas
The retrosubicular area is not directly associated with a specific Brodmann area, as it is part of the hippocampal formation, a medial temporal lobe structure not included in Brodmann's original cytoarchitectonic maps.Location
The retrosubicular area, or presubiculum, is situated in the medial temporal lobe between the subiculum and parasubiculum, forming part of the hippocampal formation (Amaral & Witter, 1995).Connections
The retrosubicular area connects with various brain regions, including the entorhinal cortex, other hippocampal subregions (e.g., subiculum, CA1), and the mammillary bodies via the fornix (Witter et al., 2000).Participation in brain networks
The retrosubicular area is part of the medial temporal lobe memory system and the Papez circuit, which are involved in memory processing, spatial navigation, and emotional processing (Aggleton & Brown, 1999; Eichenbaum, 2000).Functions
The retrosubicular area involves various functions, including spatial navigation, memory, and emotional processing (Eichenbaum, 2000).Role in clinical disorders
Abnormalities in the retrosubicular area have been implicated in several neurological disorders, such as Alzheimer's disease, temporal lobe epilepsy, and schizophrenia (Du et al., 1993; Heckers et al., 1998; Hyman et al., 1984).Areas 13, 14, and 52: Parainsular Area (junction of the temporal lobe and insula)
The parainsular area is not a well-defined or widely recognized region in the human brain, and limited information is available on this specific area. Graphics © Science and Fascija/Shutterstock.com.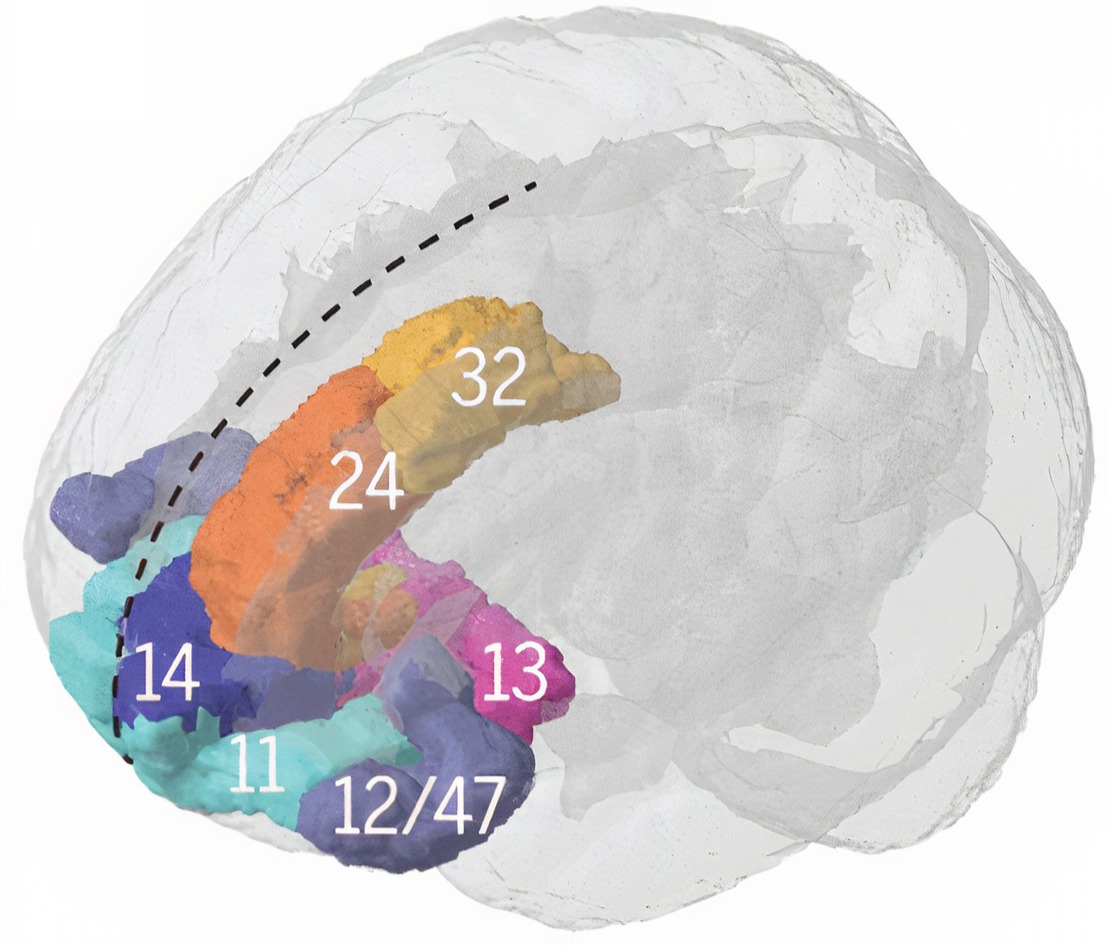
.jpg)
Brodmann areas
The insular cortex is associated with Brodmann areas 13, 14, and 52.Location
The insular cortex is located deep within the lateral sulcus, separating the frontal and parietal lobes from the temporal lobe.Connections
The insular cortex has widespread connections with various brain regions, including the prefrontal cortex, parietal cortex, temporal cortex, and limbic structures (Augustine, 1996).Participation in brain networks
The insular cortex is involved in multiple brain networks, including the salience network, and is responsible for detecting and responding to salient stimuli (Menon & Uddin, 2010).Functions
The insular cortex involves various functions, including interoception, emotional processing, pain perception, and cognitive control (Craig, 2009).Role in clinical disorders
Abnormalities in the insular cortex have been implicated in several neurological and psychiatric disorders, such as anxiety, depression, autism, and schizophrenia (Menon, 2011).Brodmann Area Involvement in Clinical Disorders
The clinical correlations between psychiatric and neurological disorders and specific Brodmann areas provide valuable insights into the neural mechanisms of these conditions. Understanding the abnormalities in these regions can offer critical information for diagnosis, treatment, and the development of targeted therapies. This section explores the associations between specific Brodmann areas and disorders including ADHD, autism spectrum disorder, bipolar disorder, major depressive disorder, panic disorder, PTSD, schizophrenia, and substance use disorder.
Attention-Deficit Hyperactivity Disorder (ADHD)
ADHD is associated with abnormalities in multiple Brodmann areas, including BAs 44/45 (Broca's area), 8/9, 10, 11, 46 (frontal regions), 7, 39, 40 (parietal regions), 4 (motor cortex), 30 (cingulate gyrus), 21, 38 (temporal regions), and 6 (premotor cortex). These abnormalities contribute to the diverse cognitive, attentional, and motor deficits observed in individuals with ADHD.
In individuals with ADHD, Broca's area (BA 44 and 45) exhibits functional disturbances, particularly a lower fractional amplitude of low-frequency fluctuations (fALFF). This dysfunction is linked to language deficits, with different ADHD subtypes affecting distinct parts of Broca's area: Area 44 in the inattentive subtype and Area 45 in the combined subtype (Pikusa & Jończyk, 2015; Silk et al., 2005).
In the frontal regions, children with ADHD display grey matter deficits in the right superior frontal gyrus (BA 8 and 9), suggesting disruptions in attentional networks (Overmeyer et al., 2001). Adolescents with ADHD show decreased activation in the middle frontal gyrus (Area 10) and the dorsolateral prefrontal cortex (Area 46), indicating widespread frontal lobe dysfunction (Silk et al., 2005). Abnormalities in the ventromedial orbitofrontal cortex (Area 11) are linked to motivational deficits in adults with ADHD (Farré-Colomés et al., 2021).
The parietal regions, including the superior parietal lobe (Area 7) and the inferior parietal lobule (Areas 39 and 40), also show decreased activation in ADHD, implicating these areas in attentional and action-attentional systems (Silk et al., 2005). Children with ADHD exhibit decreased activation in the primary motor cortex (Area 4) during motor tasks, suggesting anomalies in motor development (Gaddis et al., 2015; Gilbert et al., 2011, 2019).
The right posterior cingulate gyrus (Area 30) shows grey matter deficits in children with ADHD, which may contribute to difficulties in attentional control (Overmeyer et al., 2001). Structural alterations are also observed in the temporal regions, with lower cortical thickness in the fusiform gyrus and temporal pole (Areas 21 and 38) in children with ADHD (Hoogman et al., 2019; Karalok et al., 2019; Lake et al., 2019; McLaughlin et al., 2014). Increased connectivity in the premotor cortex (Area 6) may be associated with challenges in motor planning and execution (Hoshi & Tanji, 2007; Oldehinkel et al., 2016; Sörös et al., 2019; Suskauer et al., 2008).
Autism Spectrum Disorder (ASD)
Research indicates that abnormalities in Brodmann areas 24, 44, 45, and 10 are associated with Autism Spectrum Disorder. These abnormalities include changes in neuron size and density, gray matter volume, and functional connectivity, which are linked to the social and communication deficits characteristic of ASD. In the anterior cingulate cortex (BA 24), there is a notable reduction in cell size and cell packing density, specifically in areas 24b and 24c, along with elevated levels of glial fibrillary acidic protein (GFAP) in the white matter, indicating increased astrocyte activation. During verbal memory tasks, decreased glucose metabolism is observed in BA24, and dysregulated DNA methylation patterns in this area impact genes related to immune functions and synaptic membranes.
In the inferior frontal cortex (BA 44 and 45), there is a decrease in the size of pyramidal neurons, which play crucial roles in language processing and social behaviors (Jacot-Descombes et al., 2012). Reduced gray matter volume in the pars opercularis (BA44) and pars triangularis (BA45) is linked to difficulties in social communication (Yamasaki et al., 2010).
The anterior prefrontal cortex (BA 10) exhibits abnormal functional connectivity between the right and left hemispheres, correlating with the severity of social deficits (Kikuchi et al., 2013). Similar to BA24, BA10 also shows dysregulated DNA methylation affecting genes involved in immune response and synaptic function (Nardone et al., 2014).
Other areas affected include the medial/cingulate regions (BAs 32, 24, and 25), where lower glucose metabolism is observed during verbal memory tasks (Deery et al., 2022; Hazlett et al., 2004, 2010). There is also a reduction in white matter volumes in the posterior frontal lobe and along the cingulate arch (Aalst et al., 2021; Gardener et al., 2016).
Bipolar Disorder (BD)
Research indicates that abnormalities in several Brodmann areas, including BAs 9, 24, 38, 41, 42, 46, and 10, are associated with bipolar disorder. These abnormalities span structural changes such as reduced glial numbers and cortical volume, as well as functional and molecular disruptions including altered gene expression and DNA methylation. In BA 9 (part of the dorsolateral prefrontal cortex), abnormalities in DNA methylation and gene expression indicate its role in the pathophysiology of BD, and this area shows decreased numbers and density of glial cells, suggesting disruptions in neurochemical regulation.
The subgenual prefrontal cortex (BA 24) exhibits reduced volume and fewer glial cells, associated with familial forms of BD. This area is significant for mood regulation and reflects neurobiological changes linked to the disorder (Öngür et al., 1998; Scarr et al., 2019).
In the temporal pole (BA 38), notable differences in DNA methylation are associated with BD, indicating involvement in the molecular mechanisms underlying the disorder (Ho et al., 2019). In the primary auditory cortex (BA 41 and 42), decreased BOLD signals in response to auditory stimuli have been identified as potential biomarkers for BD (Okamoto et al., 2022).
BA 46, another region of the dorsolateral prefrontal cortex, shows altered gene expression with specific genes exhibiting differential expression in BD, suggesting disruptions in neuronal network functions (Nakatani et al., 2006; Vizueta et al., 2012). The prefrontal cortex (BA 10) also presents changes in gene expression in BD, highlighting complex regional-specific changes in cortical gene expression (Scarr et al., 2019; Vizueta et al., 2012).
Major Depressive Disorder (MDD)
Research consistently implicates several Brodmann areas in the pathology of major depressive disorder, including the anterior cingulate cortex (BA 24 and BA 32), dorsolateral prefrontal cortex (BA 9 and BA 46), ventromedial prefrontal cortex (BA 10), orbitofrontal cortex (BA 13 and BA 47/12), and subgenual cingulate cortex (BA 25). In the anterior cingulate cortex, structural abnormalities and reduced volume in the subgenual part of BA 24 are linked to familial forms of MDD, and elevated levels of tumor necrosis factor (TNF) have been observed in BA 24 among MDD patients, suggesting the involvement of pro-inflammatory pathways. Abnormalities in BA 32 have been associated with antidepressant treatment.
In the dorsolateral prefrontal cortex (BA 9 and 46), both structural and functional abnormalities have been connected to MDD. In BA 9, changes in connectivity and increased fractional amplitude of low-frequency fluctuation (fALFF) are noted (Gao et al., 2021; Lai & Wu, 2015; Ye et al., 2012; Vasic et al. 2008; Zhukovsky et al., 2020). Elevated TNF levels in BA 46 indicate a role in the disorder's pathophysiology (Dean et al., 2010).
The ventromedial prefrontal cortex (BA 10) shows abnormal functional connectivity linked to emotional regulation deficits in MDD patients (Almeida et al., 2011; Johnstone et al., 2007; Wackerhagen et al., 2017; Young et al., 2016). Epigenetic modifications, such as methylation changes, have been identified in BA 9 and BA 38 (Ho et al., 2019).
In the orbitofrontal cortex (BA 13 and BA 47/12), reduced functional connectivity in BA 13 is associated with depressive symptoms, while increased functional connectivity in BA 47/12 correlates with negative self-perception in MDD (Cheng et al., 2016). The subgenual cingulate cortex (BA 25) exhibits reduced functional connectivity with regions involved in emotional regulation (Peng et al., 2020), and methylation changes in BA 25 have been consistently found in independent brain samples (Åberg et al., 2018).
Panic Disorder
Research indicates that abnormalities in Brodmann areas 11, 25, 32, and 15, as well as regions within the prefrontal cortex and parahippocampal gyrus, are associated with panic disorder. During panic attacks, there is a noticeable decrease in regional cerebral blood flow (rCBF) in the right orbitofrontal cortex (BA 11) and the prelimbic cortex (BA 25). This reduction is also observed in the anterior cingulate cortex (BA 32), which exhibits mixed patterns of hyperactivation and hypoactivation during emotional processing.
The parahippocampal gyrus in panic disorder patients shows significant abnormal asymmetry in cerebral blood flow. The anterior temporal cortex (BA 15) experiences decreased rCBF during panic attacks (Hasler et al., 2007). Key regions within the prefronto-limbic network, particularly areas within the PFC, show selective deficits in emotional processing among panic disorder patients (Hasler et al., 2007; Shang et al., 2014). Dysfunctional communication within frontotemporal structures is indicated by decreased coherence imaging values (Shang et al., 2014; Speer et al., 2003).
Increased resting-state functional connectivity between the thalamus and insula suggests excessive sensitivity to external information in panic disorder patients, highlighting a potential neural mechanism underlying their heightened responsiveness to stimuli (Feldker et al., 2016, 2019; Zhou et al., 2022).
Post-Traumatic Stress Disorder (PTSD)
PTSD is associated with abnormalities in the anterior cingulate cortex (BA 32 and BA 24), medial prefrontal cortex (BA 10 and BA 11), dorsolateral prefrontal cortex (BA 46), insula (BA 13), orbitofrontal cortex (BA 25), and sensorimotor areas (BA 4/6). These regions are involved in emotional regulation, memory processing, and response to trauma-related stimuli. In individuals with PTSD, the DLPFC (BA 46) shows decreased activity and signs of mitochondrial dysfunction, and the anterior cingulate gyrus (BA 32) exhibits less activation compared to those without PTSD.
The medial prefrontal cortex (BA 10 and 11) shows reduced activation in PTSD subjects (Etkin & Wager, 2007; Herringa et al., 2012; Manthey et al., 2021; Sartory et al., 2013). The activation levels in BA 10 correlate with PTSD symptoms during threat processing. The insula (BA 13) demonstrates increased delta slow waves linked to worsening PTSD symptoms over time (Harricharan et al., 2019; Herringa et al., 2012; Rabinak et al., 2011), and differences in functional connectivity in the insula are observed between PTSD and non-PTSD individuals (Rabinak et al., 2011; Sripada et al., 2012; Zhang et al., 2016).
During symptom provocation in PTSD patients, there is increased regional cerebral blood flow (rCBF) in the right sensorimotor areas (BA 4 and 6), while lowered rCBF is observed in the right retrosplenial cortex (BA 26, 29, and 30) (Pissiota et al., 2000). Structural and functional changes in the hippocampus and amygdala are consistently observed in PTSD, highlighting their crucial roles in memory and emotional processing (Chen et al., 2018; Hull, 2002).
Schizophrenia
Abnormalities in frontal (BAs 4, 6, 8, 9, 10, 44, 46, 47), temporal (BAs 20, 21, 22, 37, 39, 42), and cingulate (BAs 24, 25, 29, 30, 31) cortices are associated with schizophrenia. These include disrupted intercorrelations, altered receptor binding, gene expression changes, and structural deficits. Significant reductions in gray matter volume have been observed in the left anterior hippocampus-amygdala, left parahippocampal gyrus, and left superior temporal gyrus, with correlations between the volume of the left posterior superior temporal gyrus and the severity of thought disorders.
Disruptions in D2 dopamine receptor patterns have been identified in the perirhinal, superior, and inferior temporal cortices (BA 20, 22, 37, 39, and 42). These disruptions are distinct to the temporal lobe and are not attributable to long-term antipsychotic treatment, suggesting a specific role in auditory hallucinations and other positive symptoms (Goldsmith et al., 1997; Joyce et al., 1997).
In schizotypal personality disorder, lower fractional anisotropy (FA) in the left temporal lobe (BA 20, 21, and 22) indicates alterations in white matter microstructure (Chan et al., 2018; Ellison-Wright & Bullmore, 2009; Hazlett et al., 2011; Lee et al., 2016; Szeszko et al., 2008). Schizophrenia is also associated with a thinner cortex and reduced surface area in the temporal lobe, with the most significant cortical thickness reductions at the temporal pole (Kaur et al., 2020; McCarley et al., 1999; Shenton et al., 1992). Progressive volumetric changes over time, particularly decreased temporal white matter volume, indicate ongoing structural abnormalities (Kaur et al., 2020; Mathew et al., 2014; Olabi et al., 2011).
Enhanced functional connectivity between the left DLPFC and the left mid-posterior temporal lobe has been identified in patients experiencing their first episode of schizophrenia (Zhou et al., 2007), a connectivity pattern that may contribute to the cognitive and perceptual disturbances characteristic of the disorder.
Substance Use Disorder (SUD)
Abnormalities in specific Brodmann areas, particularly BA 10, BA 24, BA 30, BA 18, BA 21/22, and BA 19, are associated with substance use disorder. These areas are involved in critical functions such as self-reflection, emotional regulation, memory, and executive function, which are often impaired in individuals with SUD.
In adolescents with SUD, there is reduced activity in BA 10 (superior, medial, and middle frontal gyrus), suggesting difficulties in self-referential evaluation and future planning (Dalwani et al., 2014). The anterior cingulate cortex (BA 24) shows abnormal activity, impacting emotional behavior and executive function in individuals with SUD.
Conclusion
The examination of specific Brodmann areas reveals significant correlations with various psychiatric and neurological disorders, underscoring the importance of these brain regions in disease pathology. ADHD features disruptions in regions related to attention and executive function. Autism spectrum disorder is characterized by changes in areas affecting social and communication skills. Bipolar disorder involves alterations in areas associated with mood regulation.
Panic disorder and PTSD both involve the prefronto-limbic network, impacting emotional and stress responses. Schizophrenia is associated with widespread disruptions affecting cognition and perception, and substance use disorder involves regions critical for executive function and emotional regulation. For biofeedback practitioners, these Brodmann area findings provide a neuroanatomical framework for understanding the EEG patterns and connectivity abnormalities observed in clinical assessment.
Key Takeaways
Brodmann areas are regions of cortex defined by their cytoarchitecture, the organization, density, and layering of neurons, first mapped by Korbinian Brodmann using Nissl staining and numbered 1 through 52, of which roughly 43 to 47 were delineated in humans. Several areas map cleanly onto function, including area 4 for primary motor cortex, area 17 for primary visual cortex, areas 44 and 45 for Broca's area, and area 22 for Wernicke's area. Many psychiatric and neurological disorders correlate with distinctive Brodmann-area abnormalities that span frontal, cingulate, temporal, and parietal regions. Probabilistic atlases such as the Jülich Brain Atlas add population-level variability that Brodmann's static map could not capture. Clinicians should treat Brodmann areas as a shared reference language while recognizing that a given area may not occupy identical cortical territory in every client.
Check Your Understanding
- What anatomical property defines a Brodmann area, and what staining method did Brodmann use to identify these regions?
- Identify the function associated with Brodmann areas 4, 17, 44 and 45, and 22.
- Give two examples of disorders discussed in this section and one Brodmann-area abnormality associated with each.
- What limitation of Brodmann's original map do modern probabilistic atlases help address?
Connectivity, Phase, and Coherence
This final section introduces the connectivity, phase, and coherence concepts that underlie EEG network analysis. These measures are central to quantitative EEG (qEEG) assessment and connectivity-based neurofeedback protocols. We will cover neural network connectivity, phase reset mechanisms, EEG synchrony, coherence measurement, and co-modulation.
Neural networks are systems of interconnected ensembles of neurons that collaborate to achieve a goal (Thompson & Thompson, 2015). Networks communicate and perform functions via hub- or node-based communication systems. The image below shows dorsal and lateral views of the connectivity backbone of the human brain, with nodes (individual ROIs) coded according to strength and edges coded according to connection weight.
Hagmann, P., Cammoun, L., Gigandet, X., Meuli, R., Honey, C. J., Wedeen, V. J., & Sporns, O. (2008) Mapping the structural core of human cerebral cortex. PLoS Biology, 6(7), e159, Wikimedia Commons, licensed under the Creative Commons Attribution 3.0 Unported license.
Connectivity
Networks like the Affect, Attention, Default, Executive, and Salience systems synchronize the activity of cortical and subcortical regions to perform functions. Connectivity is the degree of synchrony between the oscillations of specialized brain regions (nodes) within a network (Bastos & Schoffelen, 2016). Strongly connected brain regions are called hubs, and hubs can be primarily connected to nodes (vertices) within their local modules (sets of interconnected nodes) or to nodes in more distant modules (Bullmore & Sporns, 2009).
Neurofeedback training can increase or decrease connectivity using a normative database. For example, BrainMaster's BrainAvatar software allows clinicians to train specific networks, like the Default Mode Network (DMN).
See the Assumptions unit for an in-depth discussion of the Affective, Default Mode, Executive, Motor, Network, Oculomotor, Salience, and Social Networks.
Phase Reset Coordination of Neural Networks
Phase refers to the degree to which the peaks and valleys of EEG waveforms coincide. Phase measures the time shift between EEG activity in two brain regions and represents the number of elements in a network times the delay in that network. Phase graphic © petrroudny43/Shutterstock.com.
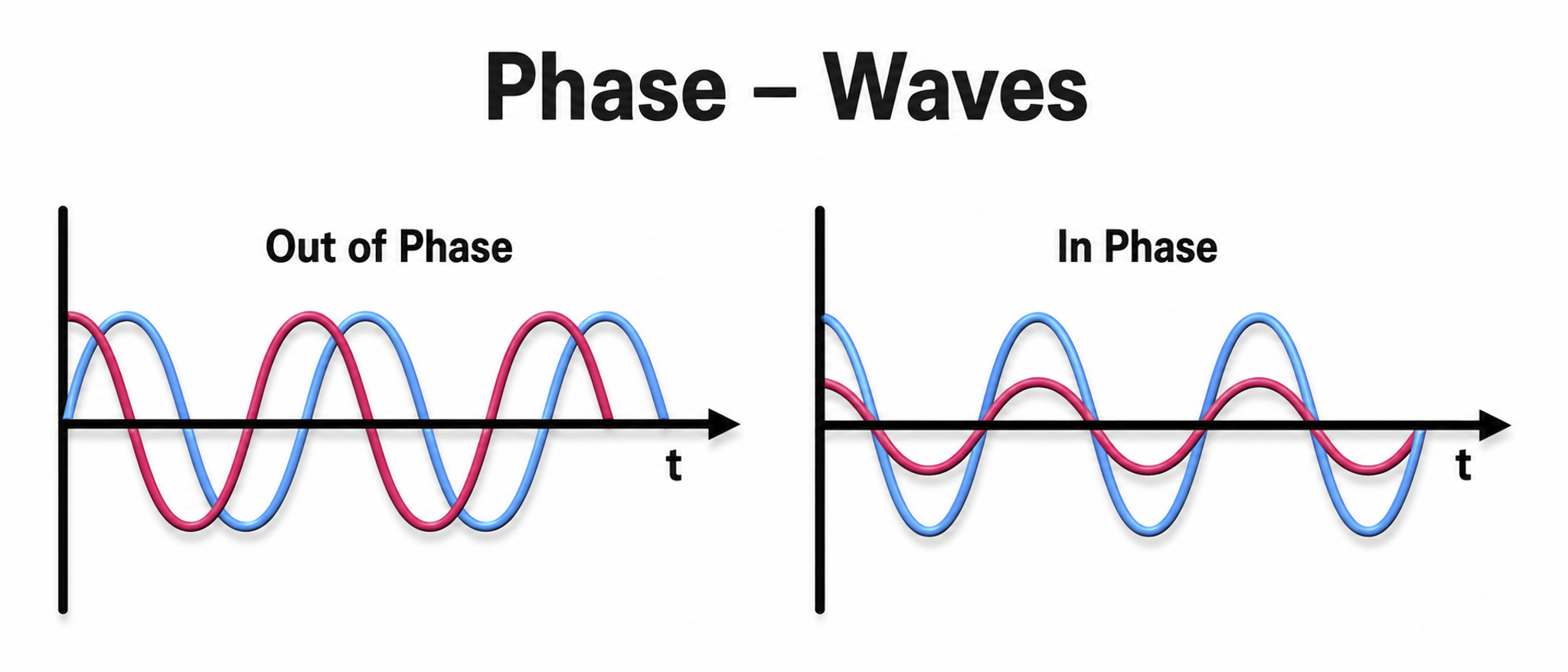
Phase reset (PR) is defined by a sudden change in phase difference (phase shift duration or SD) followed by a period of phase locking (lock duration or LD), expressed as PR = SD + LD (Thatcher et al., 2009). Gap junction coupling explains both phase locking and phase shifting (Hughes & Crunelli, 2007).
Resetting the phase of ongoing oscillatory activity to endogenous (internal) or exogenous (environmental) cues facilitates coordinated information transfer within circuits and between distributed brain areas. Phase resetting is a critical marker of dynamic state changes within functional networks (Voloh & Womelsdorf, 2016). Phase resets create a neural context, a narrow band of frequencies that uniquely characterize the activated circuits, and impose coherent low-frequency phases to which high-frequency activations can synchronize.
These identifiable cross-frequency correlations span large distances and are critical for neural coding models that depend on phase, increasing the informational content of neural representations. Phase resets likely originate from the dynamics of canonical excitatory-inhibitory (E-I) circuits that are anatomically ubiquitous. They reorganize oscillations in diverse task contexts: attentional stimulus selection, classical conditioning, cross-modal integration, sensory perception, and spatial navigation, and can drive changes in ensemble organization, functional networks, neural excitability, and overt behavior.
The frequency graphic © Ali DM/Shutterstock.com. Low- and high-frequency oscillations result from phase reset decreases and increases in neural activity, respectively.
EEG Synchrony and Phase
Networks of neurons generating the EEG activity at different sites can produce signals that are identical in amplitude, frequency, and phase, or that are entirely unrelated. Understanding the distinctions between different types of synchrony is essential for interpreting qEEG connectivity measures.
Synchrony means that the firing of pools of neurons is coordinated. EEG signals can display three types of synchrony. Local synchrony occurs when the coordinated firing of cortical neurons produces high-amplitude EEG signals. For example, an alpha amplitude of 20-60 μV detected at O1-A1 is produced by the synchronous firing of pools of neurons, while beta activity at the same site is lower in amplitude due to desynchronized firing. This is analogous to a choir: when performers sing in unison, they produce a louder sound than when singing separately.
Frequency synchrony occurs when identical EEG frequencies are detected at two or more electrode sites, for example, 12 Hz simultaneously detected at O1-A1 and O2-A2. Phase synchrony reflects the degree of alignment of oscillatory activity across regions. If the peaks of theta waves occur at the same instant at two cortical regions, then there is zero phase lag between them.
The phase alignment of two EEG signals of the same frequency reflects the speed of information transmission, with lower phase lag (greater synchrony) reflecting faster communication between sites. High synchrony within frequency bands, such as alpha or beta, supports efficient communication, while cross-frequency synchrony between different EEG bands plays a role in the integration of brain functions.
However, increased phase synchrony does not always indicate active communication between two regions. In some cases, it simply reflects two areas receiving input from the same rhythmic generators. Increased phase synchrony can also result from damage to interactive communication, particularly with delta activity in TBI, where damaged regions independently produce delta oscillations due to a lack of other input, sometimes with increased synchrony.
EEG training may produce phase-synchronous 12-Hz alpha waves at O1-A1 and O2-A2.
Coherence
Coherence represents the degree of coupling between separate cortical regions and reflects neural network connectivity and dynamics (Bullmore & Sporns, 2009). Coherence evaluates the linear association or correlation between EEG waveforms recorded from two different scalp locations using a two-channel referential montage, as shown below.
Coherence measures the degree to which two areas have consistent phase relationships at a designated frequency. When the phase difference between two signals is constant, coherence = 1; when the phase difference is random, coherence = 0.
Hyper-coherence means too much coupling, involving a failure to activate cortical regions selectively, and may interfere with multitasking and rapid decision-making. Hypo-coherence, which often results from traumatic brain injuries, means too little coupling and involves a breakdown in communication between regions that should normally coordinate (Wilson et al., 2011).
Coherence is a correlation coefficient (squared) that estimates relative amplitude and phase consistency between any pair of signals in each frequency band (Bendat & Piersol, 2010). The key component is consistency: coherence is based on phase, and phase measures the timing relationship of two waveforms at a specific EEG frequency band.
The image below shows five identical wave patterns shifted in time relative to each other. Starting with the first waveform at the bottom, each subsequent wave is shifted in time, known as phase shift or phase angle. The angle is represented in degrees: a wave beginning one-fourth of the way through the original wave's cycle has a 90-degree phase angle (example B), while one beginning at the halfway point has a 180-degree phase angle, meaning the waves are exactly opposite (example C).

As previously noted, coherence measures the consistency of these phase relationships. Waveforms do not have to be in phase (zero phase angle) to be coherent; they only need to maintain a consistent phase relationship. Coherence values range from 0 to 1, with zero representing a random relationship and 1 representing signals that remain in the same phase relationship for the entire measurement period (usually a minimum of 2 seconds).
Because signals can be out of phase and still highly coherent, coherence should not be confused with phase synchrony, which describes two waves oscillating with the same timing (zero phase angle). This distinction matters clinically.
Key Concept
Coherence and phase synchrony are related but distinct. Coherence measures the consistency of the phase relationship between two signals over time and ranges from zero, for a random relationship, to one, for a perfectly consistent one. Two waveforms can be out of phase yet highly coherent, as long as their phase difference stays constant. Phase synchrony, by contrast, describes waves oscillating with the same timing, that is, a zero phase angle. Confusing the two can lead a clinician to misread connectivity findings.
In heart rate variability (HRV) training, for example, a common goal is to synchronize respiration and heart rate waveforms. This is an example of phase synchrony. When this alignment is consistent, the two waves are also highly coherent. However, a true coherence calculation could show high coherence even when waveforms are significantly offset, as long as they remain in a consistent phase relationship.
In practice, many commercial HRV displays report a correlation- or spectrum-based index of relative phase synchrony rather than the full coherence calculation, which combines the cross-spectral density and the auto-spectral density between two signals. Practitioners should check what a given instrument actually computes before calling its output coherence.
EEG coherence estimates interaction between neural systems through amplitude and phase consistency in each frequency band between two scalp sensor locations. Two signals can be asynchronous but have high coherence if they are consistently in the same asynchronous relationship, implying some coordinating mechanism between the two locations related to communication and interaction (Nunez & Srinivasan, 2006).
The history of coherence measures in EEG began with Donald Walter (Walter, 1963), and since then many studies have used EEG coherence to identify factors in cognition, brain maturation, heritability, gender differences, and a variety of clinical disorders (Thatcher, 2012). There are several methods for estimating coherence, and each has important limitations that clinicians must understand.
Thatcher (2012) states that the linked ears montage or a common reference is necessary to correctly calculate phase differences because other montage arrangements distort the original time series and the relationships between individual sensor locations. Thatcher (2004) claims that the reliability of coherence measures using common average reference or Laplacian montages, though generally quite high, is irrelevant because these montaging methods render the resulting calculations uninterpretable. He argues that these montages involve the "mixing" of signals from multiple electrodes and provides examples using artificially-generated signals showing that only the linked-ear or common reference correctly represents changes in source signals as noise increases.
However, the linked ear and common references in Thatcher's study were necessarily neutral, without signal content, and therefore do not reflect real-world conditions. On the other hand, Nunez and Srinivasan (2006) demonstrate that coherence using a common reference (such as linked-ear or linked-mastoid sensors) elevates all coherence measures for all frequencies and electrode combinations because this method adds a common signal to each channel.
The idea of a "neutral" reference without any EEG or other electrical activity is not valid in real-world recordings. Ear and mastoid reference channels frequently show rhythmic alpha activity, particularly in the eyes-closed condition, and are often contaminated by EMG artifacts from masseter muscle contractions. Similarly, a common reference such as Cz or Fcz would add real EEG signal characteristics to the resulting calculation, influencing phase and coherence calculations, particularly for frontal electrode locations.
For a complete discussion of differential amplifier functions, please see the Instrumentation and Electronics section. For our purposes, it is sufficient to understand the concept of common-mode rejection, which eliminates common or similar signals between two inputs (positive and negative). The initial purpose was to eliminate "mains" artifact (60 Hz in North America, 50 Hz elsewhere), but common-mode rejection also removes any electrical signal, including EEG, from active/reference pairs when the signals are identical in frequency and, to a lesser extent, amplitude.
When the reference channel derived from ear or mastoid sites contains substantial alpha activity and is compared to frontal "active" channels that generally do not contain substantial alpha, the resulting signal retains the alpha from the reference. This creates the appearance of rhythmic, highly synchronous frontal alpha activity and produces artificially elevated alpha coherence values. Compared to a normative database, these values will generate falsely elevated z-scores, and neurofeedback training may be implemented to "correct" what is actually an artifact of the reference choice.
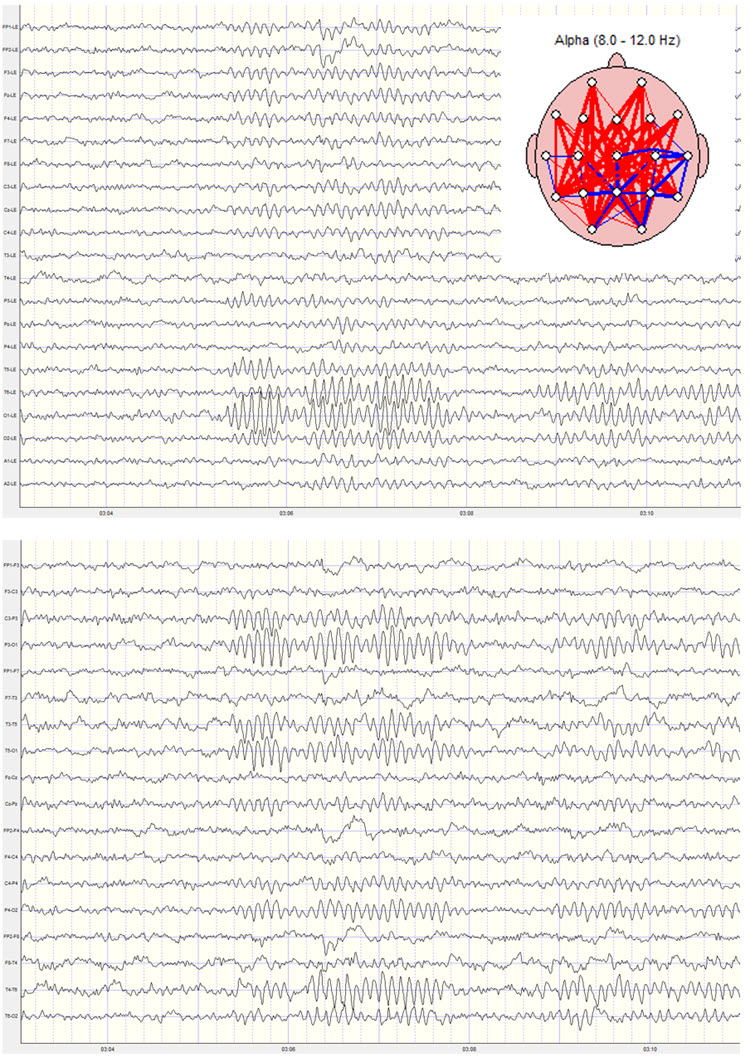
The same epoch is shown twice: as raw traces in a linked-ears referential montage (upper panel, note the A1-LE and A2-LE channels) and in a bipolar montage (lower panel), with an alpha-band connectivity head map inset at the upper right. A burst of alpha rises to prominence over the posterior channels in both montages.
The head map is labeled Alpha (8.0-12.0 Hz), and each line joins a pair of electrodes whose coherence departs from the normative mean: red marks pairs above the mean and blue marks pairs below it. The map therefore reports how consistently two sites hold a phase relationship over time. It does not report how much alpha either site produces, and it does not report whether their amplitudes rise and fall together, which is co-modulation.
Read alongside the discussion above, the dense red lattice reaching well into the frontal channels is exactly the pattern a linked-ear reference can manufacture when the ear channels themselves carry alpha. It should be confirmed in a second montage before it is treated as a clinical finding.
Nunez and Srinivasan (2006) suggest that the common average reference method shows coherence results more similar to experimental, reference-independent calculations, though they note that many sensors (their example used 111) are needed for an effective average reference, far more than the typical 19 used in most normative databases.
Another approach is the surface Laplacian method, which removes some issues associated with reference contamination though it tends to increase noise at each electrode location (Nunez & Srinivasan, 2006). It also reduces the problem of volume conduction, the conduction of bioelectric signals through neural tissues that appear in adjacent sensor locations not directly above the signal generator. Volume conduction decreases with distance, so closely spaced electrode pairs show higher coherence disproportionately affected by common sources. Widely spaced electrode pairs more accurately reflect actual communication between areas. Because the Laplacian operator uses the average of surrounding electrodes as the reference, common signals are minimized and coherence may be more accurate for closely spaced pairs.
The eventual answer to which coherence measure is better must await further studies using real-world models. Current practice with either method will produce results that require an understanding of the benefits and limitations of each approach. Excess frontal alpha coherence derived from linked ear reference data should be reviewed cautiously, and the raw EEG signal should always be viewed using multiple montages to determine the accuracy of frontal alpha displays.
Other factors influencing coherence include pervasive EMG artifacts that may not be evident to beginning practitioners. When such artifacts contaminate the record, excessive hypercoherence in beta and fast beta frequencies can result, as shown in the example below.
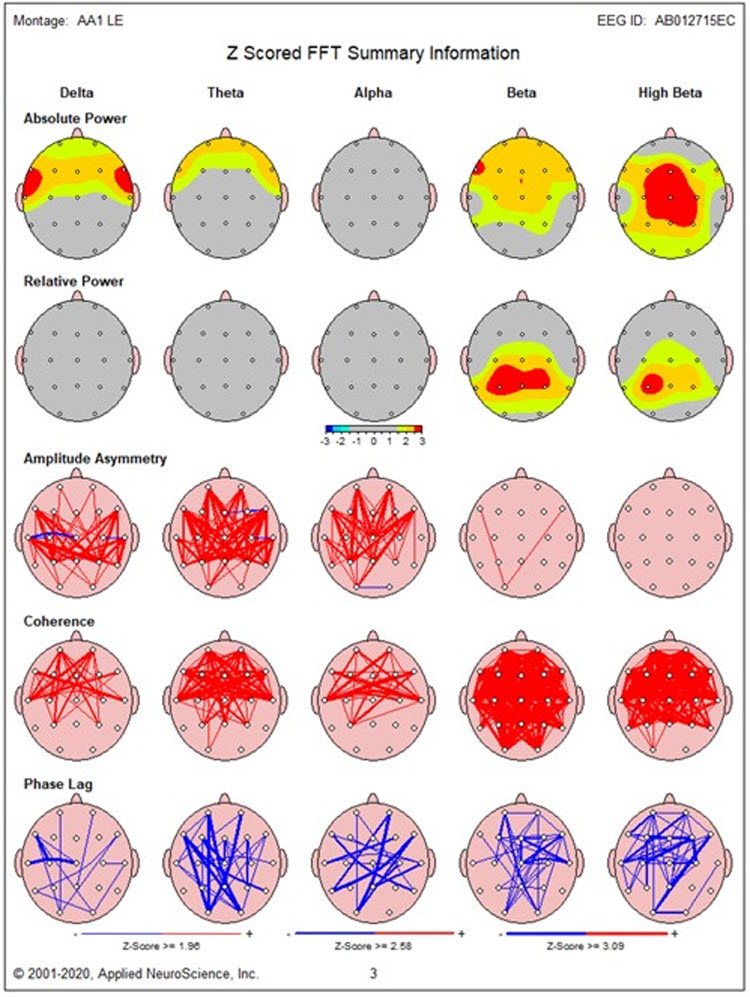
A Z-scored FFT summary recorded in a linked-ears montage (AA1 LE). Five columns run across the frequency bands from delta to high beta, and five rows step through absolute power, relative power, amplitude asymmetry, coherence, and phase lag, so reading across a row shows one measure at every band and reading down a column shows one band across every measure.
No cell shows raw microvolts. Every value is a Z score, the distance from a normative mean expressed in standard deviations, with warm colors above the mean, cool colors below it, and gray marking the unremarkable middle. The top two rows are power maps. The lower three are pairwise measures drawn as webs between electrode pairs; coherence and phase lag are connectivity measures, while amplitude asymmetry compares power at two sites rather than their coupling. The thresholds printed beneath the connectivity rows, Z of at least 1.96, 2.58, and 3.09, determine which pairs are drawn at all.
Note the combination the text above describes: elevated frontal and temporal high-beta power together with a dense red coherence lattice in beta and high beta. That pairing is a common signature of EMG contamination rather than of cortical connectivity, and the raw record should be inspected in multiple montages before these deviations are read as findings.
With these cautions in mind, coherence remains a useful tool that can be quite revealing, particularly in cases of traumatic brain injury, stroke, lesion, and other evaluations. With experience, clinicians can identify which coherence values represent actual findings and correlate them with client history and presenting symptoms.
Simple phase measurements alone are not particularly useful because they are not directional; they simply show the timing relationship between two sensor locations without revealing which location may be dysfunctional or what the causal factors may be (Nunez & Srinivasan, 2006). Additionally, a common reference distorts phase in the same way described for coherence. Nunez (1974) argues that using a bipolar montage for phase calculation substantially reduces problems of volume conduction and reference contamination, though this is not suggested as a viable montage for subsequent coherence calculation.
Phase values range from -1 to +1, and statistical calculations can be made based on normative samples. However, experimental data (Nunez, 1995; Silberstein et al., 1993) show that only about 8% of phase data could be identified as associated with traveling waves; the rest was too complex to identify source relationships. More recent work has examined phase slope, which provides a measure of effective connectivity, the directionality of influence between two EEG sources.
Co-Modulation
Co-modulation is the degree of association in the magnitude of signals detected from two sources (sites). Co-modulation, which can be measured using the Pearson Product-Moment Correlation Coefficient, shows the degree to which signals strengthen and weaken in a correlated manner (Collura, 2014). Co-modulation does not measure phase or coherence, although it may reflect phase effects (Sterman & Kaiser, 2001). It represents a complementary approach to understanding brain connectivity that avoids some of the reference-related limitations of traditional coherence measures.
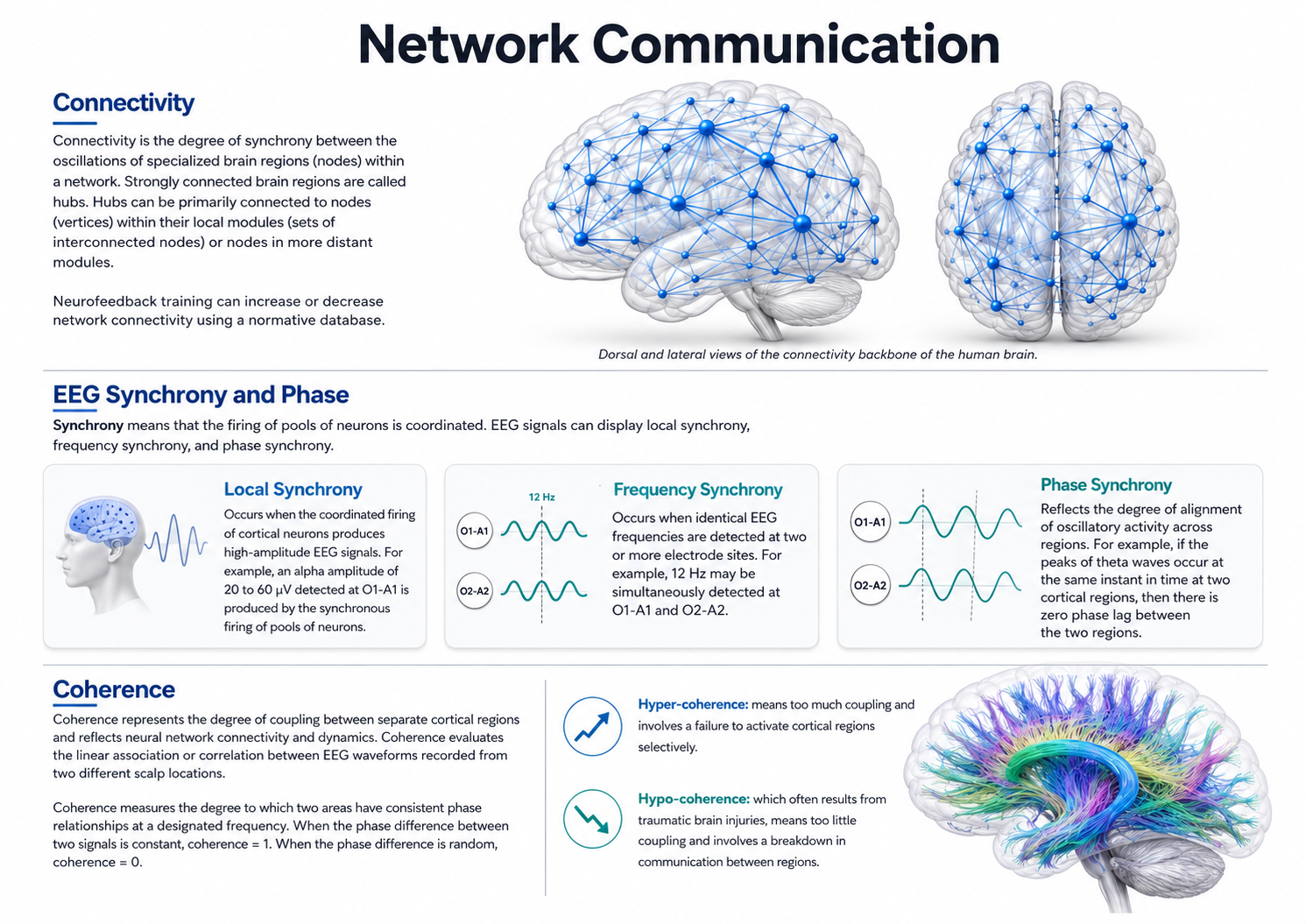
In summary, connectivity, phase, and coherence are powerful tools for understanding brain network dynamics, but each measure has specific limitations that clinicians must understand. Reference montage selection, volume conduction, and EMG artifacts can all distort these measures. Effective clinical practice requires viewing raw EEG data with multiple montages and interpreting connectivity findings in the context of client history and symptoms.
Clinical Application
A client's qEEG shows strikingly elevated frontal alpha coherence recorded with a linked-ear reference. Before designing a protocol to correct it, recall that ear and mastoid channels often carry rhythmic alpha, especially with eyes closed, and that a common reference adds that signal to every channel. The result can be artificially high frontal alpha coherence and falsely elevated z-scores against a normative database. The safeguard is to view the raw EEG in multiple montages and confirm the finding is real before training away what may be an artifact of the reference choice.
Key Takeaways
Connectivity is the degree of synchrony among specialized regions, or nodes, that are organized into hubs and modules within networks such as the Default Mode and Salience systems. Phase measures the timing shift between regions, and phase reset, expressed as shift duration plus lock duration, coordinates information transfer across circuits. Coherence measures the consistency of the phase relationship on a scale from zero to one and differs from phase synchrony, which requires a zero phase lag. Reference-montage choice, volume conduction, and EMG artifact can all distort these measures, as when a linked-ear reference produces falsely elevated frontal alpha coherence. Sound interpretation requires viewing the raw EEG in multiple montages and reading connectivity findings in the context of client history and symptoms.
Check Your Understanding
- Define connectivity, and explain the difference between a hub and a node.
- Write the equation for phase reset and define its two components.
- How does coherence differ from phase synchrony, and can two waveforms be out of phase yet highly coherent?
- Explain how a linked-ear reference can create artificially elevated frontal alpha coherence, and what a clinician should do before training to correct it.
Cutting-Edge Topics in Neuroanatomy
Meningeal Lymphatics and Glymphatic Clearance
The discovery that the dural venous sinuses house meningeal lymphatic vessels overturned the long-held belief that the central nervous system lacks lymphatic drainage (Aspelund et al., 2015; Louveau et al., 2015). Working together with the glymphatic system, which clears wastes such as amyloid-beta, tau, and alpha-synuclein mainly during slow-wave sleep, this anatomy links cerebral venous drainage to waste clearance and immune surveillance. For biofeedback practitioners, it reinforces why sleep quality belongs in every client assessment.
The Neurovascular Unit and Blood-Brain Barrier
Rather than a passive wall of endothelial cells, the blood-brain barrier is now understood as a coordinated neurovascular unit that also includes pericytes, astrocyte endfeet, microglia, and the surrounding basement membrane (Ding et al., 2025; Abbott et al., 2010). This tightly regulated selectivity protects neural tissue but excludes most small-molecule and nearly all large-molecule therapeutics, which makes the barrier a central obstacle in treating neurological and psychiatric disorders (Daneman & Prat, 2015).
Bidirectional Hippocampal Traveling Waves
Recordings from patients undergoing brain surgery show that hippocampal brainwaves travel in both directions, from back to front and from front to back, rather than in a single direction (Kleen et al., 2021). Cognitive activity influences the direction of movement differently for low- and high-frequency waveforms, refining older one-directional models of how the hippocampus integrates information.
Locus Coeruleus, Pupillometry, and Attention
Although the locus coeruleus cannot be monitored noninvasively, its activity correlates with pupil dilation, which gives researchers an indirect window on noradrenergic arousal (Dahl et al., 2020; Dahl et al., 2022). Greater alpha blocking and pupil dilation predict better performance on demanding attention tasks, connecting a small brainstem nucleus to the alpha dynamics that clinicians train.
Glossary
acetylcholine: an amine neurotransmitter that binds to nicotinic and muscarinic ACh receptors.
acetylcholine esterase (AChE): the enzyme that deactivates ACh.
AChE-R: an abnormal form of acetylcholine esterase (AChE), which may render dendrites with acetylcholine receptors more excitable when stressed.
action potential: a propagated electrical signal that usually starts at a neuron’s axon hillock and travels to presynaptic axon terminals.
adenylate cyclase: at a metabotropic receptor, an enzyme that transforms ATP into the second messenger cyclic AMP.
affective network: a network that is triggered when we make mistakes and that monitors cognitive activity to predict when errors are likely and greater executive control may be needed. It includes the anterior cingulate cortex, hippocampal cortex, entorhinal cortex, superior temporal gyrus, inferior temporal gyrus, posterior parietal cortex, globus pallidus internal segment, substantia nigra pars reticulata, and medial dorsal nucleus of the thalamus.
afferent: a neuron that transmits sensory information towards the central nervous system, or from one region to another.
all-or-none law: once an action potential is triggered in an axon, it is propagated, without decrement, to the end of the axon. The amplitude of the action potential is unrelated to the intensity of the stimulus that triggers it.
alpha blocking: arousal and specific forms of cognitive activity may reduce alpha amplitude or eliminate it entirely while increasing EEG power in the beta range.
alpha rhythm: 8-12-Hz activity that depends on the interaction between rhythmic burst firing by a subset of thalamocortical (TC) neurons linked by gap junctions and rhythmic inhibition by widely distributed reticular nucleus neurons. Researchers have correlated the alpha rhythm with "relaxed wakefulness." Alpha is the dominant rhythm in adults and is located posteriorly. The alpha rhythm may be divided into alpha 1 (8-10 Hz) and alpha 2 (10-12 Hz).
alpha spindles: regular bursts of alpha activity.
alpha-subunit: a subunit of a G protein associated with the neuron membrane that breaks away to activate enzymes within the neuron when a ligand binds to a metabotropic receptor.
amino acid neurotransmitters: the oldest family of transmitters. These molecules bind to ionotropic and metabotropic receptors, transmitting information and modulating neuronal activity. Most synaptic communication is accomplished in the brain by glutamate (generally excitatory) and GABA (generally inhibitory).
AMPA (glutamate) receptors: ionotropic receptors which open sodium channels, depolarizing the neuron's membrane (producing an EPSP), and dislodging a Mg+ ion that blocks an adjacent NMDA (glutamate) receptor's calcium channel. AMPA receptors are responsible for most activity at glutamatergic synapses.
amplitude: the size of the voltage fluctuation in an EEG signal, measured in microvolts (μV). Amplitude is not the same quantity as power, which is proportional to the square of amplitude and is expressed in μV².
amygdala: a limbic system structure that participates in evaluating whether stimuli are salient (rewarding or threatening), learning about the consequences of our actions, establishing unconscious emotional memories and conditioned emotional responses, creating declarative memories for events with emotional significance, and producing anxiety and fear responses. It is a major subcortical target of the orbitofrontal, cingulate, entorhinal, and insular cortices.
angular gyrus: located in the parietal lobe near the junction of the temporal and occipital lobes, the angular gyrus corresponds to Brodmann area 39. It plays a role in language processing, attention, spatial cognition, and integration of sensory information.
anion: negative ion, for example, chloride (Cl-).
anisotropic attenuation: the variation in the degree of signal attenuation depending on the direction of the electrical currents. This phenomenon occurs due to the heterogeneous nature of the skull's structure.
anterior: near or toward the front of the head, for example, the anterior cingulate.
anterior cingulate cortex (ACC): a medial frontal region lying above the corpus callosum (Fpz and Fz) that encompasses Brodmann areas 24, 25, 32, and 33. The ACC plays a vital role in attention, executive function, emotional regulation, conflict monitoring, and error detection, and it is activated during working memory. The ACC mediates emotional and physical pain and has cognitive (dorsal anterior cingulate) and affective (ventral anterior cingulate) conflict-monitoring components.
anterior commissure: a bundle of nerve fibers that crosses the midline and connects the left and right temporal lobes, hippocampus, and amygdala.
anterior prefrontal cortex (APFC): found in the most anterior region of the prefrontal cortex, the anterior prefrontal cortex corresponds primarily to Brodmann area 10 and extends into area 11. It supports complex cognitive processes such as planning, decision-making, working memory, prospective memory, and abstract reasoning.
apical dendrite: a dendrite that arises from the top of the pyramid and extends vertically to layer 1 of the neocortex.
arousal: a process that combines alertness and wakefulness, produced by at least five neurotransmitters, including acetylcholine, histamine, hypocretin, norepinephrine, and serotonin.
astrocytes: star-shaped glial cells that communicate with and support neurons and help determine whether synapses will form.
asynchronous waves: voltages produced when neurons depolarize and hyperpolarize independently.
ATP: the energy source for a neuron’s sodium-potassium transporters.
Attention-Deficit Hyperactivity Disorder (ADHD): a neurodevelopmental disorder characterized by inattention, hyperactivity, and impulsivity. It is associated with abnormalities in brain regions such as the prefrontal cortex and basal ganglia.
auditory cortex: temporal cortex situated in the superior temporal gyrus that processes and interprets auditory information within the dorsal and ventral streams. It comprises the primary auditory cortex (Brodmann areas 41 and 42) and the surrounding secondary auditory cortex (Brodmann area 22).
autism spectrum disorder (ASD): a neurodevelopmental disorder characterized by deficits in social communication and interaction, along with restricted and repetitive behaviors. It involves atypical brain development and connectivity, particularly in areas such as the temporal lobes and frontal cortex.
autonomic nervous system: a subdivision of the peripheral nervous system that innervates glands and internal organ smooth muscles and includes enteric, parasympathetic, and sympathetic divisions.
autoreceptors: metabotropic receptors that can be located on the membrane of any part of a neuron. They detect the neurotransmitters the neuron itself releases, generate IPSPs that inhibit the neuron from reaching the excitation threshold, and regulate internal processes like transmitter synthesis and release through the second messenger system.
axoaxonic synapses: junctions between two axons that do not affect the generation of an action potential, only the amount of neurotransmitter distributed.
axodendritic synapses: junctions between axons and dendrites determine whether the axon hillock will initiate an action potential.
axon: long, cylindrical structures that convey information from the soma to the terminal buttons. An axon also transports molecules in both directions along the outer surface of protein bundles called microtubules.
axon hillock: a swelling in the cell body where a neuron integrates the messages it has received from other neurons and decides whether to fire an action potential.
axonal varicosity: a swelling in an axon wall allowing neurotransmitter release through the wall via volume transmission.
axoplasmic transport: the movement of molecules in both directions along the outer surface of protein bundles called microtubules.
basal dendrite: dendrite that horizontally branches out from the 30-μm base of the pyramid through the layer where the neuron resides.
basal forebrain: a cholinergic network located in the ventral frontal lobe and anterior hypothalamus that influences cerebral blood flow and cognitive activity.
basal ganglia: these forebrain structures consist of an egg-shaped nucleus that contains the putamen and globus pallidus and a tail-shaped structure called the caudate, which together are responsible for the production of movement. The basal ganglia have also been implicated in obsessive-compulsive disorder, Parkinson’s disease, and Huntington’s chorea.
benzodiazepine receptor agonist (BZRA) hypnotics: nonbenzodiazepines like zolpidem (Ambien).
beta rhythm: 12-38-Hz activity associated with arousal and attention generated by brainstem mesencephalic reticular stimulation that depolarizes neurons in both the thalamus and cortex. The beta rhythm can be divided into multiple ranges: beta 1 (12-15 Hz), beta 2 (15-18 Hz), beta 3 (18-25 Hz), and beta 4 (25-38 Hz).
bilateral synchronous slow waves: a pathological sign observed in drowsy children. When detected in alert adults, intermittent bursts of high amplitude slow waves may signify gray matter lesions in deep midline structures.
bipolar disorder (BD): a mental health disorder characterized by extreme mood swings, including manic and depressive episodes. It is associated with abnormalities in brain regions involved in mood regulation, such as the prefrontal cortex and amygdala.
brain connectivity studies: research focused on understanding how different regions of the brain communicate with each other. These studies often use EEG data to map functional connections and require accurate signal interpretation to account for the effects of the skull and other tissues.
Broca's area: area located inferior frontal gyrus (BA 44 and 45) of the dominant hemisphere (F7-T3 in the left hemisphere) concerned with speech production, grammar, language comprehension, and sequencing.
Brodmann areas: cytoarchitectural zones of the cerebral cortex mapped by Korbinian Brodmann using Nissl staining and defined by their distinct cellular composition and organization. They are numbered 1 through 52, of which roughly 43 to 47 were delineated in the human cortex.
cancellous bone: also known as trabecular or spongy bone, it is found inside bones and is less dense and more porous than cortical bone. It offers slightly less resistance to electrical signals than cortical bone.
cation: a positive ion, for example, sodium (Na+).
caudal: away from the front of the head.
cell body or soma: contains machinery for cell life processes and receives and integrates EPSPs and IPSPs from axons generated by axosomatic synapses (junctions between axons and somas). The cell body of a typical neuron is 20 μm in diameter, and its spherical nucleus, which contains chromosomes comprised of DNA, is 5-10 μm across.
central autonomic network (CAN): a system of brain regions that regulates the autonomic nervous system, including the prefrontal cortex, anterior cingulate cortex, insula, amygdala, hypothalamus, periaqueductal gray, parabrachial complex, nucleus of the solitary tract, and medulla oblongata. These structures work together to regulate physiological states such as heart rate, blood pressure, respiration, digestion, and thermoregulation.
central executive network: a network including the dorsolateral prefrontal cortex, anterior cingulate cortex, and orbitofrontal cortex that is responsible for the cognitive regulation of behavior.
central nervous system (CNS): the division of the nervous system that includes the brain, spinal cord, and retina.
central nucleus of the amygdala: the nucleus that orchestrates the nervous system's response to essential stimuli by activating circuits in the brainstem (autonomic arousal) and the basal ganglia and periaqueductal gray (defensive behavior).
central sulcus: the sulcus that separates the frontal and parietal lobes, and with them the primary motor cortex from the primary somatosensory cortex.
cerebral cortex: the layer of gray matter that covers the cerebral hemispheres. The cerebral cortex consists of gray matter and white matter.
cerebral ventricles: a network of fluid-filled chambers that protects the brain from trauma due to abrupt head movements and facilitates the exchange of nutrients and wastes between blood vessels and the brain.
cerebrospinal fluid (CSF): fluid produced by the choroid plexus membrane of the lateral ventricles that fills the ventricular system.
chemical synapses: junctions between neurons that transmit molecules across gaps of roughly 200-500 angstroms (20-50 nm). Neurons use chemical synapses to produce short-duration (milliseconds) and long-duration (seconds to hours) changes in the nervous system. Chemical synapses are capable of more extensive communication and initiate more diverse and long-lasting changes than electrical synapses.
choroid plexus: a network of specialized capillaries and ependymal cells lining the brain's ventricles that produces cerebrospinal fluid and forms the blood-CSF barrier.
cingulate cortex: a part of the limbic system situated in the medial aspects of the frontal and parietal lobes. It is divided into an anterior cingulate cortex (Brodmann areas 24, 32, and 33) and a posterior cingulate cortex (Brodmann areas 23, 29, 30, and 31), and it participates in emotion processing, memory, attention, and cognitive control.
circle of Willis: vascular network located at the base of the brain comprised of the carotid and basilar arteries. This structure may provide another route for delivering blood when a major artery is compromised by disease or traumatic injury.
classical routes for EEG activation: specific sensory pathways like the visual (retina to the visual cortex), auditory (cochlea to the auditory cortex), and somatosensory (chemoreceptors and mechanoreceptors to the somatosensory cortex) systems. Increased transmission of information through these pathways desynchronizes EEG activity in the cortical regions to which these afferent neurons project, as specialized circuits of neurons independently process this information.
closed field: the extracellular field produced by neurons whose dendrites radiate symmetrically in all directions, such as many stellate and thalamocortical cells. Opposing transmembrane currents largely cancel, so these neurons contribute little to the scalp EEG.
co-modulation: the degree of association in the magnitude of signals detected from two different sources (sites). Co-modulation, which can be measured using the Pearson Product-Moment Correlation Coefficient, shows the degree to which signals strengthen and weaken in a correlated manner.
coherence: the degree of coupling between separate cortical regions and reflects neural network connectivity and dynamics. Coherence evaluates the linear association or correlation between the EEG waveforms recorded from two different scalp locations using a two-channel referential montage.
commissures: axon tracts. The left and right hemispheres communicate using the corpus callosum, anterior commissure, and posterior commissure.
complex: a sequence of waves.
COMT: a degrading enzyme that only targets the catecholamines dopamine and norepinephrine.
connectivity: the degree of synchrony between the oscillations of specialized brain regions (nodes) within a network.
contingent negative variation (CNV): a steady, negative shift in potential (15 μV in young adults) detected at the vertex. This slow cortical potential may reflect expectancy, motivation, intention to act, or attention. The CNV appears 200-400 ms after a warning signal (S1) and, with short S1-S2 intervals, peaks within 400-900 ms; the peak latency is paradigm-dependent and with longer intervals the late CNV peaks just before S2. It declines sharply after a second stimulus that requires the performance of a response (S2).
continuous irregular delta: slow waves produced by white matter lesions seen in disorders like multiple sclerosis.
contralateral: structures that are located on opposite sides of the body. For example, neurons in the left primary motor cortex control muscles on the right side of the body.
coronal plane: the plane that separates the body into front and back parts.
corpus callosum: the largest commissure that connects the left and right frontal, parietal, and occipital lobes.
cortical bone: the dense, outer surface layer of bone that provides strength and rigidity. It is one of the two types of bone found in the skull and has high resistance to electrical signals.
corticothalamic network: a unified network that generates diverse types of brain rhythms grouped by slow cortical oscillations.
cranial nerves: 12 pairs of nerves connected to the brain and are part of the sensory and motor systems of the head and neck.
cranium: the part of the skull that encloses the brain, composed of several bones fused together, including the frontal, parietal, temporal, and occipital bones.
cyclic AMP: a second messenger that moves about the neuron, activating other enzymes. Protein kinase A, which controls the excitability of ion channels, is a crucial enzyme target of cyclic AMP. Cyclic AMP also travels to the nucleus to regulate gene expression.
cytoarchitecture: the microscopic appearance and organization of neurons within the cortex, including cell types, cell densities, and the arrangement of cortical layers.
D-serine: a neurotransmitter that binds to the glycine site on the NMDA receptor to trigger calcium entry into a dendritic spine when glutamate binds to its site, resulting in a large, prolonged increase in intracellular calcium.
Dale's law: the superseded view that a neuron can release only one neurotransmitter. Many neurons release more than one.
default mode network (DMN): frontal, temporal, and parietal lobe circuits that are active during introspection and daydreaming and relatively inactive when we pursue external goals. Its cortical hubs include the ventral posterior cingulate cortex, precuneus, medial prefrontal cortex, and angular gyrus.
delta rhythm: 1-4-Hz oscillations generated by thalamocortical neurons during stage 3 (N3) sleep. The distinct slow oscillation below 1 Hz originates in the neocortex.
dendrite: a branched structure designed to receive messages from other neurons via axodendritic synapses (junctions between axons and dendrites), determining whether the axon hillock will initiate an action potential.
dendritic spines: protrusions on the dendrite shaft where axons typically form axodendritic synapses.
dendrodendritic synapses: junctions between dendrites that communicate chemically across synapses and electrically across gap junctions.
depolarize: to make the membrane potential more positive by making the inside of the neuron more positive with respect to its outside.
desynchronization: the absence or loss of coordinated neuronal firing and synchronization of brain waves.
diencephalon: the posterior forebrain subdivision that contains the thalamus and hypothalamus.
diffusion: the distribution of molecules from areas of high concentration to low concentration.
diphasic wave: a wave that contains both a negative and positive deflection from the baseline.
dipole: the electrical field generated between the sink (where current enters the neuron) and the source (place at the other end of the neuron where current leaves) may be located anywhere along the dendrite.
distal: toward the periphery.
dominant frequency: EEG frequency with the greatest amplitude.
dopamine: a monoamine neurotransmitter that exerts its postsynaptic effects on at least six receptors linked to G proteins. This means that dopamine functions as a neuromodulator. The two major families include D1 (D1 and D5) and D2 (D2A, D2B, D3, and D4).
dorsal: toward the upper back or head.
dorsal anterior cingulate cortex (dACC): located in the dorsal region of the anterior cingulate cortex, the dorsal anterior cingulate cortex includes Brodmann areas 24 and 32. It plays a role in cognitive control, decision-making, and conflict monitoring.
dorsal entorhinal cortex (DEC): situated in the medial temporal lobe, the dorsal entorhinal cortex is often associated with Brodmann areas 28 and 34, although the entorhinal cortex is allocortex and does not map neatly onto Brodmann's cytoarchitectonic divisions. It is involved in spatial memory and navigation.
dorsal posterior cingulate cortex (dPCC): the dorsal part of the posterior cingulate cortex, composed primarily of Brodmann areas 23 and 31. It is involved in self-referential thought, memory, and spatial awareness.
dorsal stream (auditory): the pathway from the temporal to the parietal lobes that helps spatially localize sounds.
dorsal stream (visual): the pathway from the primary visual cortex (V1) to the parietal lobe that helps to localize objects and guide movements towards them.
dorsolateral prefrontal cortex (DLPFC): the region of the middle frontal gyrus (BA 9 and 46, and parts of BA 8 and 10) that shares responsibility with cortical and subcortical networks for executive functions like abstract reasoning, cognitive flexibility, decision-making, inhibition, planning, and working memory (Miller & Cummings, 2007) and exercises the highest cortical level of motor control.
dual-action antidepressants: medications that activate 5-HT1 receptors to produce antidepressant and anxiolytic effects, while they blockade 5-HT2 (agitation, restlessness, and sexual dysfunction) and 5-HT3 (nausea, headache, and vomiting) receptors to minimize their side effects.
dural venous sinuses: venous channels enclosed between the two layers of the dura mater, the tough outer covering of the brain. Unlike ordinary veins, they have rigid walls formed by dura rather than a muscular tunica, and they lack valves. They collect deoxygenated blood from the brain and cerebral veins, along with cerebrospinal fluid absorbed through the arachnoid granulations, and channel it toward the internal jugular veins for return to systemic circulation.
ectosplenial cerebral cortex: part of the retrosplenial cortex, the ectosplenial cerebral cortex is situated within the posterior cingulate cortex and covers Brodmann area 29. It plays a role in spatial memory, navigation, and contextual processing.
EEG activity: a single wave or successive waves.
EEG power: the signal power in the EEG spectrum. Most EEG power falls within the 0-20 Hz frequency range. Power is proportional to the square of amplitude and is expressed in μV². μV² equals picowatts only if a 1-ohm reference resistance is assumed, and that assumption must be stated.
efferent: a motoneuron that transmits information towards the periphery.
electrical synapse: a symmetrical synapse where neurons communicate information bidirectionally across gap junctions between adjacent membranes using ions. Transmission across electrical synapses is nearly instantaneous, compared with the roughly 0.5-1-ms synaptic delay at chemical synapses. The rapid information transmission that characterizes electrical synapses enables large circuits of distant neurons to synchronize their activity and simultaneously fire.
electroencephalogram (EEG): the voltage difference between at least two electrodes, where at least one electrode is located on the scalp or inside the brain. The EEG is a recording of both EPSPs and IPSPs that occur primarily in dendrites in pyramidal cells located in macrocolumns, several mm in diameter, in the upper cortical layers.
electrostatic pressure: the attractive or repulsive force between ions that moves them from one region to another.
EMG inhibit: a threshold set on muscle-artifact activity (typically high-frequency EEG contaminated by EMG, often in the ~30-100+ Hz range) that blocks or withholds reward feedback whenever that activity exceeds the threshold. It discourages muscle tension, jaw clenching, and movement artifact, keeping reinforcement tied to genuine cortical activity rather than contamination.
entorhinal cortex: a structure located in the caudal region of the temporal lobe and that receives pre-processed sensory information from all modalities and reports on cognitive operations. The entorhinal cortex provides the main input to the hippocampus, is involved in memory consolidation and spatial localization, and provides input into the septohippocampal system that may generate the 4-7 Hz theta rhythm.
enzymatic deactivation: the process in which an enzyme breaks a neurotransmitter apart into inactive fragments. For example, acetylcholine transmission is ended by the enzyme acetylcholine esterase (AChE). Deactivating enzymes located in the synaptic cleft degrade a neurotransmitter molecule when it detaches from its binding site.
evoked potential: an event-related potential (ERP) elicited by external sensory stimuli (auditory, olfactory, somatosensory, and visual). An auditory evoked potential has a negative peak at 80-90 ms and a positive peak around 170 ms following stimulus onset; peak latencies differ by modality, and the pattern-reversal visual evoked potential peaks positively near 100 ms. The orienting response ("What is it?") is a sensory ERP. The N1-P2 complex in the auditory cortex of the temporal cortex reveals whether an uncommunicative person can hear a stimulus.
excitatory postsynaptic potential (EPSP): a brief positive shift in a postsynaptic neuron's potential produced when neurotransmitters bind to receptors and cause positive sodium ions to enter the cell. An EPSP pushes the neuron towards the excitation threshold when it can initiate an action potential.
executive functions: a set of cognitive processes, including working memory, flexible thinking, and self-control, that enable goal-directed behavior.
exocytosis: the process of neurotransmitter release. When an action potential arrives and depolarizes the terminal button, calcium ions enter the terminal button from the extracellular fluid. Calcium binds with clusters of protein molecules that join the vesicles with the presynaptic membrane. The clusters move apart, forming a hole through both membranes called a fusion pore, and the neurotransmitter leaves the terminal button for the synaptic cleft or extracellular fluid.
exogenous ERP: an event-related potential (ERP) elicited by external sensory stimuli (auditory, olfactory, somatosensory, and visual).
explicit learning: behavioral changes that occur with our conscious awareness that require processing by the hippocampus.
extracellular dipole layers: sheets of charge created by macrocolumns of pyramidal cells. The dipole layer lies parallel to the cortical surface while the pyramidal neurons that create it are oriented perpendicular to it, sending opposite charges towards the surface and towards the deepest of the six cortical layers.
extracellular fluid: the fluid surrounding a neuron.
facultative pacemaker theory: Andersen and Andersson's (1968) theory that thalamic neurons activate cortical neurons and thalamic inhibitory interneurons via recurrent collaterals.
fast cortical potentials: EEG rhythms that range from 0.5 Hz-100 Hz. The main frequency ranges include delta, theta, alpha, sensorimotor rhythm, and beta.
feature binding: the process of linking information to perceptual objects (linking an apple's color to its shape) that may involve the 40-Hz rhythm.
fissures: deep grooves, for example, the lateral fissure.
focal waves: EEG waves detected within a limited area of the scalp, cerebral cortex, or brain.
foramina: openings or holes in the skull that allow for the passage of nerves and blood vessels. These structures can create localized points where electrical signals might experience less attenuation.
forebrain: the anterior brain subdivision that consists of the cerebral hemispheres (telencephalon) and the thalamus and hypothalamus (diencephalon), also called the prosencephalon.
frequency: the number of cycles completed each second expressed in hertz (Hz).
frequency synchrony: when identical EEG frequencies are detected at two or more electrode sites. For example, 12 Hz may be simultaneously detected at O1-A1 and O2-A2.
frontal eye field (FEF): a premotor region in the anterior part of the middle frontal gyrus that corresponds to Brodmann area 8. It is involved in voluntary eye movement control and visual attention, and it directs gaze toward targets selected by the intraparietal sulcus.
frontal lobes: the most anterior cortical lobes of the brain (F7, F3, Fz, F8, F4) that are divided into the primary motor cortex, motor association cortex, Broca's area, and prefrontal cortex.
functional networks: sets of brain regions whose activity is correlated over time, in contrast to structural networks defined by axonal projections and pathways.
fusiform gyrus: a region on the ventral surface of the temporal and occipital lobes associated with Brodmann areas 37 and 19. It is involved in face recognition, object recognition, and the processing of color and visual form.
fusion pore: a hole through a vesicle and presynaptic membrane that allows neurotransmitters to leave the terminal button for the synaptic cleft or extracellular fluid.
G protein: a protein located inside a neuron’s membrane next to a metabotropic receptor, activated when the receptor binds a ligand. An alpha-subunit of the G protein then breaks away to perform actions within the cell.
GABA: an amino acid that is often inhibitory. GABA may be the most important inhibitory neurotransmitter in the brain. There are several types of GABA receptors, each producing inhibition differently.
gamma rhythms: a 28-80 Hz rhythm that includes the 38-42 Hz Sheer rhythm and is associated with learning and problem-solving, meditation, mental acuity, and peak brain function in children and adults.
gap junction: an electrical synapse, which is a symmetrical synapse where neurons communicate information bidirectionally across gap junctions between adjacent membranes using ions. Transmission across electrical synapses is nearly instantaneous, compared with the roughly 0.5-1-ms synaptic delay at chemical synapses. The rapid information transmission that characterizes electrical synapses enables large circuits of distant neurons to synchronize their activity and simultaneously fire.
generalized asynchronous slow waves: waves seen in sleepy children and those with elevated temperatures. This may indicate degenerative disease, dementia, encephalopathy, head injury, high fever, migraine, and Parkinson's disease in adults.
glial cells: nonneural cells that guide, insulate, and repair neurons and provide structural, nutritional, and information-processing support. Glial cells contribute to slow cortical potentials (SCPs), which are generated chiefly by pyramidal apical dendrites. Glial cells include astrocytes, microglia, oligodendrocytes, radial glial cells, and Schwann cells.
global loops: cortical macrocolumns separated by as much as 7 cm that receive shared input and fire synchronously to generate delta and theta rhythms.
glutamate: an amino acid that is often excitatory and that may be the primary excitatory neurotransmitter in the brain. Its receptors are found on the surface of almost all neurons. There are at least 13 different receptors for glutamate, 5 ionotropic and 8 metabotropic. Most presynaptic neurons in the brain excite postsynaptic neurons via ionotropic glutamate receptors in the postsynaptic membrane. Metabotropic glutamate receptors may play a regulatory function, either augmenting or suppressing the activation of ionotropic glutamate receptors.
glycine: an amino acid that is often inhibitory and has a binding site on the NMDA receptor.
gray matter: brain tissue that looks grayish brown and comprises cell bodies, dendrites, unmyelinated axons, glial cells, and capillaries.
gyrus: ridge of cortex demarcated by sulci or fissures, for example, the precentral gyrus.
hertz (Hz): unit of frequency, an abbreviation for cycles per second.
high alpha (alpha 2): 10-12-Hz alpha associated with open awareness.
high beta (beta 4): 25-38-Hz activity mostly seen in the frontal lobes and is associated with hyper-perfusion and increased glucose metabolism. High or fast beta activity may be related to peak performance and cognitive processing and related to specificity and precision in information processing. Excessive high beta is associated with alcoholism, anxiety, OCD, rumination, and worry.
hindbrain: posterior brain division that consists of the cerebellum, pons, and medulla.
hippocampus: a seahorse-shaped limbic structure. The hippocampus is required to form declarative memories and plays a vital role in emotion, navigation, spatial memory, and dampening the endocrine stress response. The hippocampus also contains leukocyte receptors, making it part of the feedback loop for immune system regulation. Hippocampal neurons and networks that include it are sources of the theta rhythm.
horizontal (transverse) plane: the plane that divides the brain into upper and lower parts.
hubs: highly centralized nodes through which other node pairs communicate; hubs allow efficient communication.
hyper-coherence: excessive coupling due to a failure to selectively activate cortical regions. Hyper-coherence may interfere with multitasking and rapid decision-making.
hyperpolarize: a negative shift in membrane potential (the inside becomes more negative with respect to the outside) due to the loss of positive ions or gain of negative ions.
hypo-coherence: deficient coupling due to a breakdown in communication between regions that should generally communicate with each other. Hypo-coherence often results from traumatic brain injuries.
impedance: a measure of the opposition that a circuit presents to the passage of a current when a voltage is applied. In the context of EEG, the skull's impedance affects the transmission of electrical signals from the brain to the electrodes on the scalp.
inferior colliculi: midbrain structures that integrate information about spatial localization and multiple sensory modalities, including somatosensory information.
inferior temporal gyrus (ITG): a region of the inferior temporal lobe that primarily includes Brodmann areas 20 and 37. It plays a role in visual object recognition and semantic memory.
inhibitory postsynaptic potential (IPSP): a brief negative shift in a postsynaptic neuron's potential produced when cations like potassium leave a neuron or anions (negative ions) like chloride enter a neuron, which hyperpolarizes the cell. An IPSP pushes the neuron away from the threshold of excitation.
insular cortex (insula): cortex that lies deep within the lateral sulcus that divides the temporal and parietal lobes, comprising Brodmann areas 13, 14, 15, 16, and parts of area 52. The insula is involved in interoception, self-awareness, emotional and autonomic responses to external stimuli, pain perception, and taste sensation, and it serves as an integrative hub for the salience network.
integration: the addition of EPSPs and IPSPs at the axon hillock. Neurons sum EPSPs and IPSPs over their surface in spatial integration and over ms of time in temporal integration to raise the membrane from its resting potential to the excitation threshold. EPSPs and IPSPs last from 15-200 ms, while action potentials occur in 1-2 ms.
internal carotid artery: a major paired artery that supplies blood to nearly two-thirds of the cerebral hemispheres.
interneurons: neurons that receive input from and distribute output to other neurons. Under the broad definition (neither primary sensory nor motor), interneurons are the great majority of neurons. Under the narrow, local-circuit definition, they are roughly 15-30% of cortical and hippocampal neurons and have short processes. Most are found in the central nervous system, but interneurons also occur in the enteric and peripheral autonomic systems. They provide the integration required for decisions, learning and memory, perception, planning, and movement.
intracellular fluid: the watery cytoplasm contained within a neuron.
ion: a charged atom or molecule with a positive or negative charge. Positive ions are called cations, and negative ions are called anions.
ionotropic receptor: receptor protein that contains a binding site for a ligand and an ion channel that opens when the neurotransmitter attaches to this site.
ipsilateral: structures that are located on the same side of the body. For example, the left olfactory bulb distributes axons to the left hemisphere.
irregular waves: successive waves that constantly alter their shape and duration.
kappa rhythm: bursts of alpha or theta and is detected over the temporal lobes of subjects during cognitive activity.
lambda waves: saw-toothed transient waves from 20-50 μV in amplitude and 100-250 ms in duration detected over the occipital cortex during wakefulness. These positive deflections are time-locked to saccadic movements and observed during visual scanning, as during reading.
lateral: to the side, away from the center, as in the lateral geniculate nucleus.
lateral geniculate nucleus (LGN): thalamic nucleus that relays visual information to the cortex.
lateral nucleus of the amygdala: a nucleus that processes sensory information and distributes it throughout the amygdala.
lateralized waves: waves that are primarily detected on one side of the scalp and may indicate pathology.
Layer III: the cortical layer that is the primary source of efferent corticocortical fibers.
Layer IV: the cortical layer that is the primary destination of thalamocortical afferents and intra-hemispheric corticocortical afferents.
Layer V: the cortical layer that is the primary origin of efferent fibers that target subcortical structures that have motor functions.
Layer VI: the cortical layer that projects cortico-thalamic efferent fibers to the thalamus, which, together with the thalamocortical afferents, creates a dynamic and reciprocal relationship between these two structures.
Layers I-III: cortical layers that receive corticocortical afferent fibers that connect the left and right hemispheres.
left dorsolateral prefrontal cortex: the division of the prefrontal cortex concerned with approach behavior and positive affect. It helps us select positive goals and organizes and implements behavior to achieve these goals.
limbic system: a poorly-defined widespread network of nuclei involved in emotion, motivation, learning, memory, and navigation. Three important limbic structures are the hippocampus, amygdala, and septal nuclei.
local loops: neighboring cortical macrocolumns that share input generate frequencies above 30 Hz in the high-beta and gamma ranges.
local synchrony: synchrony that occurs when the coordinated firing of cortical neurons produces high-amplitude EEG signals.
localized slow waves: waves that may indicate a transient ischemic attack (TIA) or stroke, migraine, mild head injury, or tumors above the tentorium. Deep lesions result in bilateral or unilateral delta.
locus coeruleus system: the noradrenergic branch of the ascending reticular activating system that projects to the thalamus, limbic system, and cerebral cortex, and contributes to wakefulness and vigilance for salient stimuli. Subnormal norepinephrine transmission may contribute to ADHD.
long-latency potentials: potentials that have extended latencies following stimulus onset, for example, P300 and N400 ERPs.
long-term depression (LTD): a persistent decrease in synaptic strength following low-frequency stimulation.
long-term potentiation (LTP): a persistent increase in synaptic strength following high-frequency stimulation.
low alpha (alpha 1): 8-10-Hz alpha below a client's peak alpha frequency when eyes are closed.
low-pass filter: a filter that allows signals with a frequency lower than a certain cutoff frequency to pass through and attenuates signals with frequencies higher than the cutoff frequency. The skull acts as a natural low-pass filter for EEG signals.
macrocolumns: circuits of cortical pyramidal neurons several millimeters in diameter that create extracellular dipole layers lying parallel to the surface of the cortex, sending opposite charges towards the surface and towards the deepest of the six layers of cortical neurons. Because the pyramidal neurons within a macrocolumn are aligned perpendicular to the cortical surface and share the same orientation, their postsynaptic potentials summate rather than cancel when the cells are active together.
Major Depressive Disorder (MDD): a psychological disorder characterized by persistent feelings of sadness, loss of interest, and other cognitive and physical symptoms. It is associated with abnormalities in brain regions such as the prefrontal cortex, amygdala, and hippocampus.
medial: toward the center of the body, away from the side. For example, the medial geniculate nucleus.
medial geniculate nucleus (MGN): thalamic nucleus that projects to several cortical auditory areas using two separate pathways. The MGN mainly relays frequency, amplitude, and binaural information to the auditory cortex in the temporal lobe.
medial prefrontal cortex: the division of the prefrontal cortex that integrates cognitive-affective information and helps control the hypothalamic–pituitary–adrenal (HPA) axis during emotional stress.
membrane potential: a neuron’s electrical charge created by a difference in ion distribution within and outside the neuron. A typical resting potential is about -70 mV (thousandths of a volt) since the inside of a resting axon is more negatively charged than the outside.
meninges: three protective layers (dura mater, pia mater, and arachnoid) that enclose the brain and spinal cord.
mesocortical neurons: dopaminergic neurons that project from the ventral tegmental area of the midbrain to the prefrontal cortex and excite prefrontal cortical neurons that control working memory, planning, and strategy preparation for problem solving. Underactivity in this pathway is associated with the negative symptoms of schizophrenia-like attentional deficits.
metabotropic receptors: include all G protein-linked receptors located on neurons, including autoreceptors. Neurotransmitters that bind to G protein-linked receptors are often called neuromodulators. Metabotropic receptors, which indirectly control the cell's operations, expend energy, and produce slower, longer-lasting, and more diverse changes than ionotropic receptors. Their effects can last several seconds, instead of milliseconds, because of the long-lived activity of G proteins and cyclic AMP.
metencephalon: the hindbrain subdivision that consists of the cerebellum and pons.
microtubules: hollow cylindrical protein bundles that are involved in axoplasmic transport.
midbrain: the middle division called the mesencephalon, which includes the inferior colliculi, superior colliculi, and substantia nigra.
middle temporal gyrus (MTG): a region of the middle temporal lobe that primarily includes Brodmann areas 21 and 39. It plays a role in language processing, semantic memory, and visual motion processing.
mirror neurons: neurons activated when we perform a movement or observe others perform the same activity. Mirror neurons may facilitate observational learning, understanding others' actions and intentions, and empathy.
modulating effects: neuromodulators like the monoamines alter the performance of diffuse networks of target neurons by indirectly controlling cellular operations when they bind to metabotropic receptors.
module: a set of interconnected nodes in a neural network.
monoamine neurotransmitters: amine neurotransmitters that include dopamine, norepinephrine, epinephrine (catecholamines), and serotonin (indoleamine). These neurotransmitters are released using volume transmission and generally have modulating effects, altering the performance of diffuse networks of target neurons.
monoamine oxidase (MAO): an enzyme that degrades and inactivates the monoamine neurotransmitters dopamine, norepinephrine, and serotonin.
monoamine oxidase inhibitors (MAOIs): antidepressant drugs that interfere with MAO's breakdown of monoamines and increase monoamine availability to treat clinical depression.
monophasic wave: either a single negative (upward) or positive (downward) deflection from baseline.
motor cortex: a subdivision of the frontal lobe located in the precentral gyrus and guides fine motor coordination (like writing).
motor ERPs: event-related potentials detected over the primary motor cortex (precentral gyrus) during movement. Their amplitude is proportional to the force and rate of skeletal muscle contraction.
motor nerves: efferent neurons that convey commands to glands, muscles, and other neurons.
movement-related potentials (MRPs): slow cortical potentials that occur at 1 second as subjects prepare for unilateral voluntary movements. MRPs are distributed bilaterally with maximum amplitude at Cz. The supplementary motor area and primary motor and somatosensory cortices primarily generate these potentials.
mu rhythm: 7-11-Hz waves resemble wickets and appear as several-second trains over central or centroparietal sites (C3 and C4).
multiple spike-and-slow-wave complex: multiple spikes associated with at least one slow wave.
muscarinic receptors: metabotropic ACh receptors that are stimulated by muscarine and blocked by atropine. Muscarinic receptors control smooth muscle and predominate in the CNS. In the CNS, muscarinic receptors help mediate learning, memory, attention, arousal, EEG, and postural control.
myelencephalon: the hindbrain subdivision that consists of the medulla.
myelinated axons: axons insulated by myelin by oligodendrocytes in the central nervous system and Schwann cells in the peripheral nervous system.
N1-P2: a sensory event-related potential in the auditory cortex of the temporal cortex that reveals whether an uncommunicative person can hear a stimulus.
N400 potential: an event-related potential (ERP) elicited when we encounter semantic violations like ending a sentence with a semantically incongruent word ("The handsome prince married the beautiful fish"), or when the second word of a pair is unrelated to the first (BATTLE/GIRL).
negative SCPs: slow cortical potentials attributed chiefly to synchronous depolarization of pyramidal apical dendrites, with a contribution from glia, that increase the probability of neuron firing.
nerve: bundled axons outside of the central nervous system.
neural network: a system of interconnected ensembles of neurons that collaborate to achieve a goal. These networks communicate and perform functions via hub- or node-based communication systems.
neuroaxis: an imaginary line that runs centrally through the central nervous system (CNS) from the front of the prefrontal cortex to the base of the spinal cord.
neuromodulator: neurochemical that modifies the effect of neurotransmitters through mechanisms like binding to metabotropic receptors.
neuron: a nerve cell that is the fundamental anatomical unit of the nervous system.
nicotinic ACh receptor: an ionotropic receptor that is stimulated by nicotine and blocked by curare. They are mainly found in the PNS on skeletal muscles. At CNS axoaxonic synapses, they produce presynaptic facilitation (increase neurotransmitter release). In the CNS, nicotinic receptors help regulate cortical blood flow, anxiety reduction, and decision-making.
nigrostriatal pathway: a dopaminergic pathway from the substantia nigra to the basal ganglia (caudate nucleus and putamen) that controls movement. The nigrostriatal pathway is progressively destroyed in Parkinson’s disease.
Nissl stain: a histological stain that binds to the rough endoplasmic reticulum of neurons, making cell bodies visible so that cortical layering and cell density can be examined.
nitric oxide: a gaseous retrograde transmitter that is involved in long-term potentiation (LTP).
NMDA (glutamate) receptors: ligand-gated and voltage-gated glutamate receptors that bind the glutamate agonist NMDA. NMDA receptors play an important role in long-term potentiation (LTP).
node: a vertex within a neural network.
nodes of Ranvier: gaps between myelinated axon segments where the axon membrane is exposed to extracellular fluid and action potentials are regenerated by sodium ion entry.
norepinephrine: a monoamine neurotransmitter that exerts postsynaptic effects at alpha and beta receptors, each with two subtypes. All norepinephrine receptors are G protein-linked. The cell bodies of the most critical noradrenergic system are located in the locus coeruleus, a nucleus found in the dorsal pons.
nucleus accumbens: a limbic structure that receives dopamine released by the mesolimbic pathway. The nucleus accumbens plays a critical role in reinforcing diverse activities, including ingestion of drugs like central nervous system stimulants.
nucleus reticularis: a thalamic nucleus that may function as a pacemaker by releasing the inhibitory transmitter GABA at synapses with thalamocortical neurons.
occipital lobes: cortical lobes (Oz, O1, O2) posterior to the parietal lobes. The primary visual cortex (V1) is located within the calcarine sulcus (BA 17). They process visual information from the eyes in collaboration with the frontal, parietal, and temporal lobes.
odd-ball stimulus: a meaningful stimulus that is different from others in a series used to elicit the P300 potential. For example, a colored playing card is presented in a series of monochrome cards.
open awareness: the ability to adaptively respond to various environmental changes.
open field: the extracellular field produced by neurons whose long dendrites are aligned in parallel and perpendicular to the cortical surface, such as cortical pyramidal neurons. Their synchronous synaptic currents create aligned dipoles whose fields summate, making these neurons the principal generators of the scalp EEG.
orbitofrontal cortex (OFC): the frontal lobe subdivision (BA 10, 11, 12, 13, and 47) that may aid planning by evaluating the consequences (rewards and punishments) of our actions and helping to generate the motivation to ingest drugs. The OFC appears to adjust decision-making based on the stakes involved and enables us to switch between substantial (investments) and trivial (snacks) choices. The OFC also supports emotional regulation and social cognition.
orienting response: Pavlov’s "What is it?" reaction to stimuli like the sound of a vase crashing that includes (1) increased sensory sensitivity, (2) head (and ear) turning toward the stimulus, (3) increased muscle tone (reduced movement), (4) EEG desynchrony, (5) peripheral constriction and cephalic vasodilation, (6) a rise in skin conductance, (7) heart rate slowing, and (8) slower, deeper breathing.
P300 potential: an event-related potential (ERP) with a latency of roughly 250-500 ms that lengthens with task difficulty, age, and cognitive impairment, and with greatest positive peaks located over parietal lobe sites. The P300 potential may reflect an event’s subjective probability, meaning, and transmission of information.
panic disorder (PD): an anxiety disorder characterized by recurrent, unexpected panic attacks and persistent concern about future attacks. It involves dysfunction in brain regions such as the amygdala and prefrontal cortex.
parahippocampal gyri: structures located within the medial temporal lobe that form spatial and nonspatial contextual associations, which serve as building blocks for contextual processing, episodic memory, navigation, and scene processing. They may also play a role in emotional responsiveness.
parainsular area: a region at the junction of the temporal lobe and insula that is associated with Brodmann areas 13, 14, and 52. It participates in auditory and somatosensory integration and in processing pain and temperature sensations.
parietal lobes: cortical lobes (Pz, P3, P4) posterior to the frontal lobes divided into the primary somatosensory cortex (postcentral gyrus) and secondary somatosensory cortex. Their primary function is to process somatosensory information like pain and touch.
pars opercularis: the opercular part of the inferior frontal gyrus, corresponding to Brodmann area 44. It plays a role in language production and forms part of Broca's area.
pars orbitalis: the orbital part of the inferior frontal gyrus, corresponding to Brodmann area 47. It is involved in language processing, social cognition, and emotional regulation.
pars triangularis: the triangular part of the inferior frontal gyrus, corresponding to Brodmann area 45. It is involved in language processing and forms part of Broca's area.
perception-action cycles: cognitive and emotional processes that adapt (and preadapt) us to our environment.
peripheral nervous system (PNS): autonomic and somatic nervous system neurons and nerves outside the skull and spinal cord.
perirhinal cortex (PRC): a medial temporal lobe region associated with Brodmann areas 35 and 36. It plays a role in object recognition, associative memory, and contextual processing.
phase: the degree to which the peaks and valleys of EEG waveforms coincide. Phase measures the time shift between EEG activity in two brain regions.
phase reset: a sudden change in phase difference (phase shift duration or SD) followed by a period of phase locking (lock duration or LD). PR = SD + LD.
phase synchrony: synchrony when identical EEG frequencies are detected at two or more electrode sites, and the peaks and valleys of the EEG waveforms coincide. This is also called global synchrony. For example, EEG training may produce phase-synchronous 12-Hz alpha waves at O1-A1 and O2-A2.
polyphasic (multiphasic) wave: a wave that contains two or more deflections of opposite polarity from baseline.
positive SCPs: slow cortical potentials attributed chiefly to pyramidal apical dendrites, with a contribution from glia, that decrease the probability of neuron firing.
Post-Traumatic Stress Disorder (PTSD): a psychological condition triggered by experiencing or witnessing a traumatic event. It involves dysfunction in brain regions such as the amygdala, hippocampus, and prefrontal cortex.
postcentral gyrus: primary somatosensory cortex, posterior to the central sulcus.
posterior: near or toward the back of the head.
posterior basic rhythm: posterior alpha rhythm associated with IQ and memory performance.
posterior cerebral arteries: the left and right posterior arterial branches of the basilar artery that supply blood to the posterior cerebral hemispheres, cerebellum, and brainstem.
posterior commissure: axon tracts located below the corpus callosum connect the right and left diencephalon and mesencephalon.
posterior cortex: parietal, temporal, and occipital cortical areas concerned with perception and memory.
precentral gyrus: primary motor cortex, anterior to the central sulcus.
prefrontal cortex (PFC): the most anterior frontal lobe division (BA 9, 10, 11, 12, 46, 47) that is subdivided into dorsolateral, medial, orbitofrontal, and anterior cingulate regions and is responsible for executive functions like attention, working memory, prediction of the outcomes of current and hypothetical actions, the ability to work toward goals, problem-solving, planning, and the ability to suppress actions that could lead to unwanted outcomes.
premotor cortex (motor association cortex): the frontal lobe subdivision (BA 6) that is anterior to the motor cortex and helps to program and execute head, trunk, and limb movements.
presynaptic facilitation: a modulatory process in which a neuron increases the presynaptic neuron's neurotransmitter release by delivering a neurotransmitter that increases calcium ion entry into its terminal button.
presynaptic inhibition: a modulatory process in which a neuron decreases neurotransmitter release by reducing calcium ion entry.
primary gustatory cortex (PGC): a region associated with Brodmann area 43, which lies at the opercular part of the inferior frontal gyrus and extends into the insular cortex (Brodmann area 13). It is responsible for processing taste information.
primary motor cortex (M1): the frontal lobe region located along the precentral gyrus (BA 4) that organizes the opposite side of the body's muscles and movements required for fine motor coordination in tasks like writing. It contains the large pyramidal neurons known as Betz cells. Lesions can result in loss of motor control, including rigid paralysis.
primary somatosensory cortex (S1): parietal lobe subdivision located in the parietal lobe's postcentral gyrus posterior to the central sulcus (BA 3, 1, and 2). S1 maps tactile and proprioceptive information, along with pain and temperature, from the opposite side of the body.
primary visual cortex (V1): the occipital lobe region located in the calcarine sulcus (BA 17, also called striate cortex) that receives most visual information from the lateral geniculate nucleus of the thalamus and performs the initial processing of visual information received from the retinas.
protein kinase A: an intracellular enzyme that controls the excitability of ion channels and is a vital enzyme target of cyclic AMP.
pyriform cortex: a ventral temporal lobe region, also called the primary olfactory cortex, that is conventionally placed at Brodmann area 27 even though it is allocortex and is not defined by Brodmann's cytoarchitectonic scheme. It is responsible for processing olfactory information.
raphe system: the midline network of cell bodies within the brainstem and midbrain that may influence alertness and vigilance through reciprocal connections with the suprachiasmatic nucleus of the hypothalamus.
rate law: the principle that neurons represent the intensity of a stimulus by variation in the rate of axon firing.
readiness potential: slow-rising, negative potential (10-15 μV) detected at the vertex before voluntary and spontaneous movement. The late component of this slow cortical potential precedes voluntary action by 0.5 to 1 s, while the early component begins roughly 1.5 to 2 s before movement onset. It peaks when the subject responds.
regional loops: cortical macrocolumns that share input and are separated by several centimeters generate alpha and beta rhythms.
regular or monomorphic waves: successive waves with identical shapes. Regular waves may resemble sine waves (sinusoidal) or maybe arched (resembling wickets), or saw-toothed (asymmetrical and triangular).
resonant loop: the synchronous firing by macrocolumns that share afferent input to generate an electrical potential.
resting potential: the membrane potential of a neuron when it is not influenced by messages from other neurons.
resting-state functional connectivity (RSFC): a neuroimaging method that identifies brain networks whose regions show synchronous activity when a person is not performing an explicit task. It is used to assess coordination between brain regions and to characterize baseline neural organization.
reticular activating system (RAS): a network of 90 nuclei within the central brainstem from the lower medulla to the upper midbrain. The reticular formation sends axons to the spinal cord, thalamus, and cortex, contributing to diverse functions like neurological reflexes, muscle tone and movement, attention, arousal, and sleep.
retinotopic organization: the mapping of spatial information from the visual field onto the surface of the visual cortex, so that neighboring points in the visual scene are represented by neighboring points in cortex.
retrosplenial cingulate cortex: a region in the posterior part of the cingulate cortex that comprises Brodmann areas 29 and 30. It is involved in spatial memory, navigation, and contextual processing.
retrosubicular area: a small medial temporal lobe region of the hippocampal formation designated Brodmann area 48, although it was not included in Brodmann's original cytoarchitectonic maps. It is involved in spatial navigation and memory.
reuptake: the primary method that neurons terminate the action of neurotransmitters. Reuptake transporters located in terminal buttons and astrocytes remove neurotransmitters from the synaptic cleft.
reward deficiency syndrome: Blum’s hypothesis that an abnormal form of the A1 allele is present in most severe alcoholics and results in defective D2 receptors. Reduced D2 receptor activity may reduce the activation of the nucleus accumbens and hypothalamus and result in dysphoria, drug craving, and compulsive drug-seeking and abuse.
rhythmic slow wave activity: a posterior waveform generated by the limbic system and thalamus that is mostly seen in the frontal-midline (FCz) when awake with eyes open.
right dorsolateral prefrontal cortex: division of the prefrontal cortex that organizes withdrawal-related behavior and negative affect and mediates threat-related vigilance. It plays a role in working memory for object location.
rostral: toward the front of the head.
sagittal plane: the plane that divides the body into right and left halves.
salience network: a network including the insula and anterior cingulate cortex that monitors the external and internal environments to determine which inputs are salient and require further processing and attention.
saltatory conduction: action potential conduction in myelinated axons in which action potentials jump from node to node, conducting roughly 10 to 100 times faster than in unmyelinated axons of comparable diameter.
secondary somatosensory cortex (S2): the region of the parietal lobe adjacent to S1 (BA 40 and 43), receives projections from it and maps touch and pain from both sides of the body.
secondary visual cortex (V2): a region adjacent to the primary visual cortex in the occipital lobe that corresponds to Brodmann areas 18 and 19. It processes visual information, including the recognition of shapes, colors, and spatial orientation.
sensorimotor rhythm (SMR): 12-15 Hz (beta 1) rhythm that is located over the sensorimotor strip (C3, Cz, C4). The waves are synchronous. SMR increases with stillness and decreases with movement. Deficient SMR may be observed in movement spectrum complaints like hyperactivity and tics.
sensorimotor system: in Sterman’s model, ascending pathways that convey information about touch and proprioception to the thalamus, the thalamus and its thalamic projections to the sensorimotor cortex, and the sensorimotor cortex, and its efferent fibers.
sensory event-related potentials (ERPs): event-related potentials evoked by external sensory stimuli (auditory, olfactory, somatosensory, and visual). For auditory stimuli, these evoked potentials or exogenous ERPs have a negative peak around 80-90 ms and a positive peak about 170 ms following stimulus onset; latencies differ by modality. They are changes in brain activity produced in response to specific stimuli. ERPs can be detected throughout the cortex. Investigators monitor ERPs by placing electrodes at locations like the midline (Fz, Cz, and Pz). A computer analyzes a subject's EEG responses to the same stimulus or task over many trials to subtract random EEG activity. Under a fixed stimulus, task, and recording site, an ERP is highly replicable in morphology, with negative and positive peaks at characteristic latencies. Morphology and topography differ across modalities, tasks, ages, and individuals, so ERPs do not always have the same waveform.
sensory nerves: neurons specialized for sensory intake. They are called afferent because they transmit sensory information towards the central nervous system.
septal nuclei: a limbic structure that contains several nuclei involved in emotion, control of aggressive behavior, reward, and addiction. The septohippocampal system contributes to the theta rhythm.
septohippocampal system: a subcortical circuit from the septum to the hippocampus that contributes to 4-7 Hz theta activity.
septum: a limbic structure that contains several nuclei involved in emotion and addiction and control of aggressive behavior.
sharp transients: a sequence that contains several sharp waves.
sharp waves: waves that resemble spikes with a pointed peak with a longer 70-200-ms duration.
sink: a site where current enters the neuron. Positive sodium ion entry into a neuron creates an active sink, represented by -ve.
sLORETA: standardized low-resolution brain electromagnetic tomography, an inverse solution that estimates the distribution of intracerebral current density from scalp-recorded EEG and reports the result in anatomical coordinates such as Brodmann areas.
slow cortical potentials (SCPs): gradual changes in the membrane potentials of cortical dendrites that last from 300 ms to several seconds. These potentials include the contingent negative variation (CNV), readiness potential, movement-related potentials (MRPs), and P300 and N400 potentials. SCPs modulate the firing rate of cortical pyramidal neurons by exciting or inhibiting their apical dendrites. They group the classical EEG rhythms using these synchronizing mechanisms.
sodium (Na+) ions: positive ions that enter a neuron during EPSPs and action potentials.
sodium-potassium transporters: pumps that are powered by ATP and that exchange three sodium for two potassium ions.
soma: see cell body or soma.
somatic nervous system: spinal nerves that innervate somatosensory receptors in the skin, joints, and skeletal muscles.
somatosensory association cortex (SAC): a posterior parietal region that encompasses Brodmann areas 5 and 7. It integrates and interprets somatosensory information such as touch, pain, temperature, and proprioception.
somatotopic organization: the mapping of body parts onto the cortical surface, in which the amount of cortex devoted to a region reflects the precision required rather than the physical size of that region.
source: the place at the end of the neuron opposite of the sink where current leaves. The source is symbolized by +ve. The extracellular area surrounding the source becomes electrically positive.
source localization: a method used in EEG analysis to determine the origin of electrical activity within the brain. Accurate source localization requires accounting for the attenuation and distortion of signals caused by the skull and other tissues.
spatial summation: the addition of EPSPs and IPSPs over a neuron’s surface.
spike: a negative transient with a pointed peak at conventional paper speeds, 20-70 ms duration, and 40-100 μV amplitude.
spike-and-slow-wave complex: a spike followed by a higher amplitude slow wave at 3 Hz. In an absence seizure, the amplitudes are very high (e.g., 160 μV).
spinal cord: the column of neurons and glial cells within the vertebral canal that extend from the brainstem to the lumbar vertebrae. The spinal cord distributes sensory information from the body to the brain, and CNS commands from the brain to the body. The spinal cord also contains networks that control reflexes and central pattern generators.
spinal nerve: 31 pairs of nerves that exit the spinal cord.
spindle waves: waves that originate in the thalamus and occur during unconsciousness and stage-2 sleep.
striatal: basal ganglia (caudate nucleus and putamen).
striate cortex: another name for Brodmann area 17, derived from the visible band of myelinated fibers in layer IV that distinguishes it from surrounding cortex.
stroke: cerebrovascular accident (CVA) involves the destruction of brain tissue (infarction) due to cerebral hemorrhage and cerebral ischemia affecting blood vessels that supply the brain. CVAs show abrupt onset and involve temporary or permanent neurological symptoms like aphasia, paralysis, or loss of sensation.
Stroop test: cognitive monitoring task where color and names conflict.
subgenual ventromedial prefrontal cortex (subgenual vmPFC): the subgenual sector of the ventromedial prefrontal cortex, consisting primarily of Brodmann areas 25 and 24b. It is involved in emotional regulation, decision-making, and social cognition.
substance use disorder (SUD): a condition characterized by the harmful or hazardous use of psychoactive substances, including alcohol and illicit drugs. It involves dysregulation in reward pathways and prefrontal cortex functioning.
substantia nigra: the midbrain structure that projects to the basal ganglia (caudate nucleus and putamen) to control movement and is progressively destroyed in Parkinson's disease.
sulcus: a shallow groove in the surface of the cerebral hemisphere, for example, the central sulcus.
superior colliculus: the dorsal midbrain structure that receives visual information and directs visual gaze and attention to selected stimuli.
superior olivary nuclei: pontine structures that process binaural information to localize sound.
superior temporal gyrus (STG): a region in the superior temporal lobe that includes Brodmann area 22 and, to some extent, areas 39 and 40. It supports auditory processing, language comprehension, and social cognition, and it contains Wernicke's area.
supplementary motor cortex (SMA): a region on the medial aspect of the superior frontal gyrus that corresponds to Brodmann area 6. It is involved in planning and coordinating complex movements and in motor learning.
supramarginal gyrus: a part of the inferior parietal lobule that corresponds to Brodmann area 40. It is involved in phonological processing, language perception, attention, spatial cognition, and the integration of sensory information.
sutures: fibrous joints that connect the bones of the skull. These joints are immovable and allow for the growth of the skull in children and adolescents.
Sylvian fissure: deep fissure that serves as the upper boundary of the temporal lobe.
synapse-associated polyribosome complexes (SPRCs): organelles with dendrites that can produce proteins that allow rapid remodeling of synapses. A polyribosome complex consists of several ribosomes bound to messenger RNA (mRNA). SPRCs represent one mechanism underlying synaptic plasticity.
synaptic cleft: 20-50 nm (200-500 angstroms) fluid-filled gap between presynaptic and postsynaptic structures.
synchronous: adverb meaning that groups of neurons depolarize and hyperpolarize simultaneously.
synchronous "alpha": network-wide "alpha" that integrates perception and facilitates action. This distributed activity appears to block localized alpha-like patterns such as mu and the posterior rhythm in favor of more broadly distributed network integration.
synchrony: the coordinated firing of pools of neurons. EEG signals can display local synchrony, frequency synchrony, and phase synchrony.
telencephalon: the frontal forebrain subdivision that consists of the cerebral cortex, basal ganglia, and limbic system.
temporal lobes: lobes separated from the rest of the cortical lobes by the Sylvian fissure, containing Brodmann areas 20, 21, 22, 37, 38, 41, and 42 laterally and 20, 27, 28, 34, 35, 36, 37, and 38 medially. Areas 39 and 40 belong to the parietal lobe. The temporal lobes process hearing, smell, and taste information and help us understand spoken language and recognize visual objects and faces.
temporal summation: the addition of EPSPs and IPSPs over time. Summation is more effective when postsynaptic potentials are generated more closely in time.
temporopolar area: the most anterior part of the temporal lobe, associated with Brodmann area 38. It is involved in olfactory processing, social cognition, and semantic memory.
terminal buttons: buds located on the ends of axon branches that form synapses and release neurochemicals to other neurons. They contain vesicles that store neurotransmitters for release when an action potential arrives. A terminal button’s presynaptic membrane may have reuptake transporters that return neurotransmitters from the synapse or extracellular space for repackaging.
thalamus: the forebrain structure above the hypothalamus that consists of specialized nuclei that process and relay data to and from the telencephalon (cerebral cortex, basal ganglia, and limbic system). The thalamus analyzes all sensory data except olfaction before distributing this information to the cortex via thalamocortical afferent fibers. The thalamus contributes to SCPs, delta, theta, alpha, SMR activity, and beta-gamma activity.
theta rhythm: 4-7-Hz rhythms, which some authors extend to 8 Hz, generated by a cholinergic septohippocampal system that receives input from the ascending reticular formation and a noncholinergic system that originates in the entorhinal cortex, which corresponds to Brodmann areas 28 and 34 at the caudal region of the temporal lobe.
threshold of excitation: the membrane potential at which an axon initiates an action potential, nominally -40 mV.
transient: a single wave or sequence of regular waves, called a complex, distinguishable from background EEG activity.
triphasic wave: a wave that contains three deflections from baseline.
unmyelinated axons: smaller-diameter axons without fatty insulation that conduct more slowly than myelinated axons.
+ve: The source is the place at the other end of the neuron where the current leaves. The source is symbolized by +ve.
-ve: A sink is where the current enters the neuron. Positive sodium ion entry into a neuron creates an active sink, symbolized by -ve.
ventral: toward the base of the skull or front of the body.
ventral anterior cingulate cortex (vACC): the ventral subdivision of the anterior cingulate cortex, comprising Brodmann areas 24 and 25 and, in some parcellations, area 33. It is involved in emotional regulation, attention, and pain processing.
ventral entorhinal cortex (vEC): a medial temporal lobe region associated with Brodmann area 28, although the entorhinal cortex is allocortex and is not readily defined by Brodmann areas. It is involved in object recognition, memory, and contextual processing.
ventral posterior cingulate cortex (vPCC): the ventral part of the posterior cingulate cortex, comprising primarily Brodmann area 23. It supports self-referential thought, episodic memory retrieval, and emotional processing.
ventral posterior nucleus (VPN): a thalamic nucleus that receives somatosensory information following crossover at the medulla and projects to the primary somatosensory cortex (S1).
ventral stream (auditory): the subcortical auditory pathway to the auditory cortex that appears to analyze sound components, including speech sounds.
ventral stream (visual): the pathway from the primary visual cortex (V1) to the inferior temporal and frontal areas that allows us to identify objects and faces.
ventral striatum: the olfactory tubercle and nucleus accumbens.
ventral tegmental area: the midbrain structure that distributes dopaminergic axons to the nucleus accumbens. Serotonin receptors on endorphin-releasing neurons in the hypothalamus may increase the activity of dopaminergic reward pathways by inhibiting the release of GABA at receptors on cell bodies of the ventral tegmental area neurons.
ventromedial prefrontal cortex (VMPFC): the ventromedial reward network (BA 10, 14, 25, 32, and parts of 11, 12, and 13) implicated in making decisions where the outcomes are uncertain and where moral values must be applied to actual situations.
vigilance system: in Sterman’s model, a system that consists of both specific brainstem nuclei (e.g., locus coeruleus and raphe nuclei) and their diffuse connections with the thalamus and other subcortical structures, and the cortex. Several neurotransmitter systems mediate vigilance, including cholinergic/glutamatergic (reticular formation), noradrenergic (locus coeruleus), and serotonergic (raphe) neurons.
visual association cortex: a set of occipital and temporal lobe regions comprising Brodmann areas 18, 19, 21, 22, and 37 that includes areas V3, V4, and V5. It is responsible for higher-level visual processing, including object recognition, motion perception, and spatial awareness.
volume conduction: the movement of the EEG through body tissues and interstitial fluid to the scalp.
volume transmission: extrasynaptic neurotransmitter release from axonal varicosities, dendrites, and terminal button into the extracellular space. Monoamines like norepinephrine and serotonin are released outside the synaptic cleft.
waveform: the shape and form of an EEG signal.
Wernicke's aphasia: a language disorder following damage to Wernicke's area, characterized by fluent but nonsensical speech and impaired comprehension of language.
Wernicke's area: area of the temporoparietal cortex (BA 22) of the dominant hemisphere specialized for the comprehension of spoken and written language. Damage can result in an inability to understand the meaning of speech and to construct meaningful sentences, while speech output remains fluent.
white matter: the layer beneath the cortex that mainly consists of myelinated axons.
Test Yourself on ClassMarker
Click on the ClassMarker logo below to take a 10-question exam over this entire unit. There is no password.

Review Flashcards on Quizlet
Click on the Quizlet logo to review our chapter flashcards.

Visit the BioSource Software Website
BioSource Software offers Physiological Psychology, which satisfies BCIA's Neuroanatomy requirement, and the Neurofeedback100 Testing Service, which provides extensive multiple-choice testing over the Neurofeedback Blueprint.

Assignment
Now that you have completed this unit, explain the importance of slow cortical potentials. How does 8-10 Hz alpha differ from 10-12 Hz alpha?
References
Aalst, J., Devrome, M., Weehaeghe, D., Rezaei, A., Radwan, A., Schramm, G., Ceccarini, J., Sunaert, S., Koole, M., & Laere, K. (2021). Regional glucose metabolic decreases with ageing are associated with microstructural white matter changes: A simultaneous PET/MR study. European Journal of Nuclear Medicine and Molecular Imaging. https://doi.org/10.1007/s00259-021-05518-6
Abbott, N. J., Patabendige, A. A. K., Dolman, D. E. M., Yusof, S. R., & Begley, D. J. (2010). Structure and function of the blood-brain barrier. Neurobiology of Disease, 37(1), 13–25. https://doi.org/10.1016/j.nbd.2009.07.030
Aberg, K., Dean, B., Shabalin, A., Chan, R., Han, L., Zhao, M., Grootheest, G., Xie, L., Milaneschi, Y., Clark, S., Turecki, G., Penninx, B., & Oord, E. (2018). Methylome-wide association findings for major depressive disorder overlap in blood and brain and replicate in independent brain samples. Molecular Psychiatry, 25, 1344-1354. https://doi.org/10.1038/s41380-018-0247-6
Advokat, C. D., Comaty, J. E., & Julien, R. M. (2019). Julien's primer of drug action: A comprehensive guide to the actions, uses, and side effects of psychoactive drugs (14th ed.). Worth Publishers.
Aggleton, J. P., O'Mara, S. M., Vann, S. D., Wright, N. F., Tsanov, M., & Erichsen, J. T. (2010). Hippocampal-anterior thalamic pathways for memory: Uncovering a network of direct and indirect actions. Eur J Neurosci, 31(12), 2292-2307. https://doi.org/10.1111/j.1460-9568.2010.07251.x
Ahuja, A., & Yusif Rodriguez, N. (2022). Is the dorsolateral prefrontal cortex actually several different brain areas? The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 42(33), 6310–6312. https://doi.org/10.1523/JNEUROSCI.0848-22.2022
Almeida, J., Kronhaus, D., Sibille, E., Langenecker, S., Versace, A., LaBarbara, E., & Phillips, M. (2011). Abnormal left-sided orbitomedial prefrontal cortical–amygdala connectivity during happy and fear face processing: A potential neural mechanism of female MDD. Frontiers in Psychiatry, 2. https://doi.org/10.3389/fpsyt.2011.00069
Aloisi, F. (2001). Immune function of microglia. Glia, 36, 165–179. https://doi.org/10.1002/glia.1106
Amaral, D. G., & Witter, M. P. (1995). Hippocampal formation. In G. Paxinos (Ed.), The rat nervous system (pp. 443-493). Academic Press.
Aminoff, E. M., Kveraga, K., & Bar, M. (2013), The role of the parahippocampal cortex in cognition. Trends Cogn Sci, 17(8), 379-390. https://doi.org/10.1016/j.tics.2013.06.009
Amunts, K., & Zilles, K. (2015). Architectonic mapping of the human brain beyond Brodmann. Neuron, 88, 1086-1107. https://doi.org/10.1016/j.neuron.2015.12.001
Amunts, K., Malikovic, A., Mohlberg, H., Schormann, T., & Zilles, K. (2000). Brodmann's Areas 17 and 18 brought into stereotaxic space, where and how variable? NeuroImage, 11, 66-84. https://doi.org/10.1006/nimg.1999.0516
Amunts, K., Schleicher, A., Bürgel, U., Mohlberg, H., Uylings, H. B., & Zilles, K. (1999). Broca's region revisited: Cytoarchitecture and intersubject variability. The Journal of Comparative Neurology, 412(2), 319–341. https://doi.org/10.1002/(sici)1096-9861(19990920)412:2<319::aid-cne10>3.0.co;2-7
Amzica, F., & Lopes da Silva, F. H. (2018). Cellular substrates of brain rhythms. In Schomer, D. L. & F. H. Lopes da Silva (Eds.). Niedermeyer's electroencephalography: Basic principles, clinical applications, and related fields (7th ed.). Oxford University Press.
Andersen, P., & Andersson, S. A. (1968). Physiological basis of the alpha rhythm. Appleton-Century-Crofts.
Andreassi, J. L. (2007). Psychophysiology: Human behavior and physiological response (5th ed.). Lawrence Erlbaum and Associates, Inc.
Ardila, A., Bernal, B., & Rosselli, M. (2015). Language and visual perception associations: Meta-analytic connectivity modeling of Brodmann area 37. Behavioural Neurology, 2015, 565871. https://doi.org/10.1155/2015/565871
Arnsten, A. F. (2006). Fundamentals of Attention-Deficit/Hyperactivity Disorder: Circuits and pathways. Journal of Clinical Psychiatry, 67 (Suppl. 8), 7-12.
Aspelund, A., Antila, S., Proulx, S. T., Karlsen, T. V., Karaman, S., Detmar, M., Wiig, H., & Alitalo, K. (2015). A dural lymphatic vascular system that drains brain interstitial fluid and macromolecules. Journal of Experimental Medicine, 212(7), 991–999. https://doi.org/10.1084/jem.20142290
Augustinack, J., Huber, K., Stevens, A., Roy, M., Frosch, M., Kouwe, A., Wald, L., Leemput, K., Mckee, A., & Fischl, B. (2013). Predicting the location of human perirhinal cortex, Brodmann's area 35, from MRI. NeuroImage, 64, 32-42. https://doi.org/10.1016/j.neuroimage.2012.08.071
Augustine J. R. (1996). Circuitry and functional aspects of the insular lobe in primates including humans. Brain Research. Brain Research Reviews, 22(3), 229–244. https://doi.org/10.1016/s0165-0173(96)00011-2
Avidan, G., & Behrmann, M. (2009). Functional MRI reveals compromised neural integrity of the face processing network in congenital prosopagnosia. Current Biology: CB, 19(13), 1146–1150. https://doi.org/10.1016/j.cub.2009.04.060
Azevedo, F. A. C., Carvalho, L. R. B., Grinberg, L. T., Farfel, J. M., Ferretti, R. E. L., Leite, R. E. P., Jacob Filho, W., Lent, R., & Herculano-Houzel, S. (2009). Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. The Journal of Comparative Neurology, 513(5), 532-541. https://doi.org/10.1002/cne.21974
Babiloni, C., Babiloni, F., Carducci, F., Cincotti, F., Del Percio, C., Hallett, M., Moretti, D. V., Romani, G. L., & Rossini, P. M. (2002). High resolution EEG of sensorimotor brain functions: Mapping ERPs or mu ERD? In R. C. Reisin, M. R. Nuwer, M. Hallett, & C. Medina (Eds.), Advances in clinical neurophysiology (Supplements to Clinical Neurophysiology Vol. 54). Elsevier Science B. V.
Bastos, A. M., & Schoffelen, J-M. (2016). A tutorial review of functional connectivity analysis methods and their interpretational pitfalls. Front Syst Neurosci. https://doi.org/10.3389/fnsys.2015.00175
Bear, M. F., Connors, B. W., & Paradiso, M. A. (2020). Neuroscience: Exploring the brain (Enhanced 4th ed.). Jones & Bartlett Learning.
Beauregard, M., & Levesque, J. (2006). Functional magnetic resonance imaging investigation of the effects of neurofeedback training on the neural bases of selective attention and response inhibition in children with Attention-Deficit/Hyperactivity Disorder. Applied Psychophysiology and Biofeedback, 31(1), 3-20. https://doi.org/10.1007/s10484-006-9001-y
Bendat, J. S., & Piersol, A. G. (2010). Random data: Analysis and measurement procedures (4th ed.). Wiley.
Birbaumer, N., Elbert, T., Canavan, A. G., & Rockstroh, B. (1990). Slow potentials of the cerebral cortex and behavior. Physiological Reviews, 70(1), 1-41. https://doi.org/10.1152/physrev.1990.70.1.1
Breedlove, S. M., & Watson, N. V. (2023). Behavioral neuroscience (10th ed.). Sinauer Associates, Inc.
Brodmann, K. (1909). Vergleichende Lokalisationslehre der Grosshirnrinde in ihren Prinzipien dargestellt auf Grund des Zellenbaues. Barth.
Broyd, S. J., Demanuele, C., Debener, S., Helps, S. K., James, C. J., & Sonuga-Barke, E. J. (2009). Default-mode brain dysfunction in mental disorders: A systematic review. Neuroscience and Biobehavioral Reviews, 33(3), 279–296. https://doi.org/10.1016/j.neubiorev.2008.09.002
Bullmore, E., & Sporns, O. (2009). Complex brain networks: Graph theoretical analysis of structural and functional systems. Nature, 10, 186-198. https://doi.org/10.1038/nrn2575
Buzsáki, G., Anastassiou, C. A., & Koch, C. (2012). The origin of extracellular fields and currents: EEG, ECoG, LFP and spikes. Nature Reviews Neuroscience, 13(6), 407-420. https://doi.org/10.1038/nrn3241
Cameron, H. A., & Dayer, A. G. (2008). New interneurons in the adult neocortex: small, sparse, but significant? Biol Psychiatry, 63(7), 650-655. https://dx.doi.org/10.1016%2Fj.biopsych.2007.09.023
Cao, Z., Ottino-Gonzalez, J., Cupertino, R. B., Schwab, N., Hoke, C., Catherine, O., Cousijn, J., Dagher, A., Foxe, J. J., Goudriaan, A. E., Hester, R., Hutchison, K., Li, C. R., London, E. D., Lorenzetti, V., Luijten, M., Martin-Santos, R., Momenan, R., Paulus, M. P., Schmaal, L., … Garavan, H. (2021). Mapping cortical and subcortical asymmetries in substance dependence: Findings from the ENIGMA Addiction Working Group. Addiction Biology, 26(5), e13010. https://doi.org/10.1111/adb.13010
Caplan, D. (2006). Why is Broca’s area involved in syntax? Cortex, 42, 469–471. https://doi.org/10.1016/S0010-9452(08)70379-4
Carlson, N. R., & Birkett, M. A. (2017). Physiology of behavior (12th ed.). Pearson Education, Inc.
Catmur, C., Walsh, V., & Heyes, C. (2007). Sensorimotor learning configures the human mirror system. Cur Biol, 17(17), 1527-1531. https://doi.org/10.1016/j.cub.2007.08.006
Chan, C. Y., Ke, D. S., & Chen, J. Y. (2009). Essential fatty acids and human brain. Acta Neurol Taiwan, 18(4), 231-241. PMID: 20329590
Collura, T. F. (2014). Technical foundations of neurofeedback. Taylor & Francis.
Costanzo, R. M. (1991). Regeneration of olfactory receptor cells. CIBA Found Symp, 160, 233-242.
Creuzfeldt, O. D. (1995). Cortex cerebri. Oxford University Press.
Cruces, R., Muñoz-García, I., Palmer-Cancel, S. J., & Salas, C. (2022). A neuropsychological rehabilitation framework to address cognitive and neurobehavioral impairments after strokes to the anterior communicating artery. Frontiers in Human Neuroscience, 16, 808011. https://doi.org/10.3389/fnhum.2022.808011
Dahl, M. J., Mather, M., & Werkle-Bergner, M. (2022). Noradrenergic modulation of rhythmic neural activity shapes selective attention. Trends in Cognitive Sciences, 26(1), 38-52. https://doi.org/10.1016/j.tics.2021.10.009
Dahl, M. J., Mather, M., Sander, M. C., & Werkle-Bergner, M. (2020). Noradrenergic responsiveness supports selective attention across the adult lifespan. The Journal of Neuroscience, 40(22), 4372-4390. https://doi.org/10.1523/JNEUROSCI.0398-19.2020
Damasio, A. (2010). Self comes to mind. Pantheon Books.
Daneman, R., & Prat, A. (2015). The blood-brain barrier. Cold Spring Harbor Perspectives in Biology, 7(1), a020412. https://doi.org/10.1101/cshperspect.a020412
Dean, B., Tawadros, N., Scarr, E., & Gibbons, A. (2010). Regionally-specific changes in levels of tumour necrosis factor in the dorsolateral prefrontal cortex obtained postmortem from subjects with major depressive disorder. Journal of Affective Disorders, 120(1-3), 245-248. https://doi.org/10.1016/j.jad.2009.04.027
deCharms, R. C., Fumiko, M., Glover, G. H., Ludlow, D., Pauly, J. M., Soneji, D., Gabrieli, J. D. E., & Mackey, S. C. (2005). Control over brain activation and pain learned by using real-time functional MRI. Proceedings of the National Academy of Sciences, 102(51), 18626-18631.
DeLong, M. R. (1990). Primate models of movement disorders of basal ganglia origin. Trends Neurosci, 13(7), 281-285. https://doi.org/10.1016/0166-2236(90)90110-v
Demos, J. N. (2019). Getting started with neurofeedback (2nd ed.). W. W. Norton & Company.
Diamond, A. (2013). Executive functions. Annu Rev Psychol, 64, 135-168. https://doi.org/10.1146/annurev-psych-113011-143750
Ding, L., Kshirsagar, P., Agrawal, P., & Murry, D. J. (2025). Crossing the blood-brain barrier: Innovations in receptor- and transporter-mediated transcytosis strategies. Pharmaceutics, 17(6), 706. https://doi.org/10.3390/pharmaceutics17060706
Dyro, F. M. (1989). The EEG handbook. Little, Brown and Company.
Eisenberger, N. I., Lieberman, M. D., & Williams, K. D. (2003). Does rejection hurt? An fMRI study of social exclusion. Science, 302, 290-292. https://doi.org/10.1126/science.1089134
El-Boustani, S., Ip, J., Breton-Provencher, V., Knott, G., Okuno, H., Bito, H., & Sur, M. (2018). Locally coordinated synaptic plasticity of visual cortex neurons in vivo. Science, 360(6395), 1349-1354. https://doi.org/10.1126/science.aao0862
Ellison-Wright, I., & Bullmore, E. (2009). Meta-analysis of diffusion tensor imaging studies in schizophrenia. Schizophrenia Research, 108, 3-10. https://doi.org/10.1016/j.schres.2008.11.021
Enticott, P. G., Kennedy, H. A., Rinehart, N. J., Tonge, B. J., Bradshaw, J. L., Taffe, J. R., Daskalakis, Z. J., & Fitzgerald, P. B. (2012). Mirror neuron activity associated with social Impairments but not age in Autism Spectrum Disorder. Biol Psychiatry, 71(5), 427-433. https://doi.org/10.1016/j.biopsych.2011.09.001
Evans, J. R., & Abarbanel, A. (1999). Introduction to quantitative EEG and neurofeedback. Academic Press.
Farré-Colomés, Á., Gerhardt, S., Luderer, M., Sobanski, E., Kiefer, F., & Vollstädt-Klein, S. (2021). Common and distinct neural connectivity in Attention Deficit/Hyperactivity Disorder and Alcohol Use Disorder: A study using resting-state functional magnetic resonance imaging. Alcoholism, Clinical and Experimental Research. https://doi.org/10.1111/acer.14593
Farwell, L. A., & Donchin, E. (1991). The truth will out: Interrogative polygraphy (“lie detection”) with event-related brain potentials. Psychophysiology, 28, 531–547. https://doi.org/10.1111/j.1469-8986.1991.tb01990.x
Fisch, B. J. (1999). Fisch and Spehlmann's EEG primer: Basic principles of digital and analog EEG (3rd ed.). Elsevier.
Fischer, D. B., Boes, A. D., Geerling, J. C., Edlow, B., Pascual-Leone, A., & Fox, M. (2016). The neuroanatomic basis of coma in humans: A study of brainstem lesions and their cortical networks (S52.003). Neurology, 84(14), Supplement S52.003. https://dx.doi.org/10.1212%2FWNL.0000000000003404
Fuster, J. (2015). The prefrontal cortex (5th ed.). Academic Press.
Garrett, B. (2003). Brain and behavior. Thompson/Wadsworth.
Hale, J. B., & Fiorello, C. A. (2004). School neuropsychology: A practitioner's handbook. Guilford Press.
Hansson, E., & Rönnbäck, L. (2003). Glial neuronal signaling in the central nervous system. FASEB J, 17, 341-348. https://doi.org/10.1096/fj.02-0429rev
He, B., & Li, G. (2010). Attenuation of EEG signals by the skull: an in vitro study of skull conductivity and its effect on dipole source localization. Medical & Biological Engineering & Computing, 48(5), 497-507.
Herbet, G., & Duffau, H. (2020). Revisiting the functional anatomy of the human brain: Toward a meta-networking theory of cerebral functions. Physiological Reviews, 100(3), 1181-1228. https://doi.org/10.1152/physrev.00033.2019
Hindriks, R., & van Putten, M. J. A. M. (2013). Thalamo-cortical mechanisms underlying changes in amplitude and frequency of human alpha oscillations. NeuroImage, 70, 150-163. https://doi.org/10.1016/j.neuroimage.2012.12.018
Hugdahl, K. (1995). Psychophysiology: The mind-body perspective. Harvard University Press.
Hughes S. W., & Crunelli, V. (2007). Just a phase they're going through: The complex interaction of intrinsic high-threshold bursting and gap junctions in the generation of thalamic alpha and theta rhythms. Int J Psychophysiol, 64(1), 3-17. https://doi.org/10.1016/j.ijpsycho.2006.08.004
Hughes, S. W., & Crunelli, V. (2005). Thalamic mechanisms of EEG alpha rhythms and their pathological implications. The Neuroscientist, 11(4), 357-372. https://doi.org/10.1177/1073858405277450
Izhikevich, E. M., & Edelman, G. M. (2008). Large-scale model of mammalian thalamocortical systems. Proceedings of the National Academy of Sciences of the United States of America, 105(9), 3593-3598. https://doi.org/10.1073/pnas.0712231105
Johnstone, T., Reekum, C., Urry, H., Kalin, N., & Davidson, R. (2007). Failure to regulate: Counterproductive recruitment of top-down prefrontal-subcortical circuitry in Major Depression. The Journal of Neuroscience, 27, 8877 - 8884. https://doi.org/10.1523/JNEUROSCI.2063-07.2007
Kalat, J. W. (2019). Biological psychology (13th ed.). Cengage Learning.
Kandel, E., Koester, J. D., Mack, S. H., & Siegelbaum, S. (2021). Principles of neural science (6th ed.). McGraw-Hill Education.
Kennerley, S. W., Behrens, T. E., & Wallis, J. D. (2011). Double dissociation of value computations in orbitofrontal and anterior cingulate neurons. Nat Neurosci, 14(12), 1581-1589. https://doi.org/10.1038/nn.2961
Kirschstein, T., & Köhling, R. (2009). What is the source of the EEG? Clinical EEG and Neuroscience, 40(3), 146-149. https://doi.org/10.1177/155005940904000305
Kitamura, T., Saitoh, Y., Takashima, N., Murayama, A., Niibori, A., Ageta, H., . . . Inokuchi, K. (2009). Adult neurogenesis modulates the hippocampus-dependent period of associative fear memory. Cell, 139(4), 814-827. https://doi.org/10.1016/j.cell.2009.10.020
Kleen, J. K., Chung, J. E., Sellers, K. K., Zhou, J., Triplett, M., Lee, K., Tooker, A., Haque, R., & Chang, E. F. (2021). Bidirectional propagation of low frequency oscillations over the human hippocampal surface. Nature Communications, 12(2764). https://doi.org/10.1038/s41467-021-22850-5
Klein, S. B., & Thorne, B. M. (2007). Biological psychology. Worth Publishers.
Krauss, G. L., Fisher, R. S., & Kaplan, P. W. (Eds.) (2011). The Johns Hopkins atlas of digital EEG: An interactive training guide. The Johns Hopkins University Press.
Kringelbach, M. L. (2005). The human orbitofrontal cortex: Linking reward to hedonic experience. Nat Rev Neurosci, 6(9), 691-702. https://doi.org/10.1038/nrn1747
Kropotov, J. D. (2009). Quantitative EEG, event-related potentials and neurotherapy. Academic Press.
Landisman, C. E., & Connors, B. W. (2005). Long-term modulation of electrical synapses in the mammalian thalamus. Science, 310(5755), 1809-1813. https://doi.org/10.1126/science.1114655
Lopes da Silva, F. (2010). EEG: Origin and measurement. In EEG - fMRI: Physiological basis, technique, and applications. Springer.
Louveau, A., Smirnov, I., Keyes, T. J., Eccles, J. D., Rouhani, S. J., Peske, J. D., Derecki, N. C., Castle, D., Mandell, J. W., Lee, K. S., Harris, T. H., & Kipnis, J. (2015). Structural and functional features of central nervous system lymphatic vessels. Nature, 523(7560), 337–341. https://doi.org/10.1038/nature14432
Lubar, J. F. (1997). Neocortical dynamics: Implications for understanding the role of neurofeedback and related techniques for the enhancement of attention. Applied Psychophysiology and Biofeedback, 22(2), 111-126. https://doi.org/10.1023/a:1026276228832
Manthey, A., Sierk, A., Brakemeier, E., Walter, H., & Daniels, J. (2021). Does trauma-focused psychotherapy change the brain? A systematic review of neural correlates of therapeutic gains in PTSD. European Journal of Psychotraumatology, 1. https://doi.org/10.1080/20008198.2021.1929025
McCarley, R., Wible, C., Frumin, M., Hirayasu, Y., Levitt, J., Fischer, I., & Shenton, M. (1999). MRI anatomy of schizophrenia. Biological Psychiatry, 45, 1099-1119. https://doi.org/10.1016/S0006-3223(99)00018-9
Menon, V., & Uddin, L. Q. (2010). Saliency, switching, attention and control: A network model of insula function. Brain Struct Funct, 214(5-6), 655-667. https://dx.doi.org/10.1007%2Fs00429-010-0262-0
Meshorer et al. (2002). Alternative splicing and neuritic mRNA translocation under long-term neuronal hypersensitivity. Science, 295(5554), 508-512. https://doi.org/10.1126/science.1066752
Miller, B. L., & Cummings, J. L. (Eds.) (2007). The human frontal lobes: Functions and disorders (2nd ed.). Guilford Press.
Molenberghs, P., Cunnington, R., & Mattingley, J. B. (2011). Brain regions with mirror properties: A meta-analysis of 125 human fMRI studies. Neurosci Biobehav Rev, 36(1), 341-349. https://doi.org/10.1016/j.neubiorev.2011.07.004
Mölle, M., Marshall, L., Gais, S., & Born, J. (2002). Grouping of spindle activity during slow oscillations in human non-rapid eye movement sleep. The Journal of Neuroscience, 22, 10941-10947. https://doi.org/10.1523/JNEUROSCI.22-24-10941.2002
Monastra, V. J., Lubar, J. F., Linden, M., VanDeusen, P., Green, G., Wing, W., . . . Fenger, T. N. (1999). Assessing attention deficit hyperactivity disorder via quantitative electroencephalography: An initial validation study. Neuropsychology, 13(3), 424-433. https://doi.org/10.1037/0894-4105.13.3.424
Monti, J. M., & Jantos, H. (2008). The roles of dopamine and serotonin, and of their receptors, in regulating sleep and waking. Prog Brain Res, 172, 625-646. https://doi.org/10.1016/S0079-6123(08)00929-1
Munro, C. A., McCaul, M. E., Wong, D. F., Oswald, L. M., Zhou, Y., Brasic, J., Kuwabara, H., Kumar, A., Alexander, M., Ye, W., & Wand, G. S. (2006). Sex differences in striatal dopamine release in healthy adults. Biological Psychiatry, 59(10), 966-974. https://doi.org/10.1016/j.biopsych.2006.01.008
Nash, J. M. (2011). The gift of mimicry. Your brain: A user's guide. Time.
Neumann, N., Strehl, U., & Birbaumer, N. (2003). A primer of electroencephalographic instrumentation. In M. Schwartz & F. Andrasik (Eds.). Biofeedback: A practitioner's guide (3rd ed.). Guilford Press.
Niedermeyer, E., & Lopes da Silva, F. H. (2004). Electroencephalography: Basic principles, clinical applications, and related fields (5th ed.). Lippincott Williams & Wilkins.
Nunez, P. (1995). Neocortical dynamics and human EEG rhythms. Oxford University Press.
Nunez, P. L., & Srinivasan, R. (2006). Electric fields of the brain: The neurophysics of EEG (2nd ed.). Oxford University Press.
Olabi, B., Ellison-Wright, I., McIntosh, A., Wood, S., Bullmore, E., & Lawrie, S. (2011). Are there progressive brain changes in schizophrenia? A meta-analysis of structural magnetic resonance imaging studies. Biological Psychiatry, 70, 88-96. https://doi.org/10.1016/j.biopsych.2011.01.032
Oldehinkel, M., Beckmann, C., Pruim, R., Oort, E., Franke, B., Hartman, C., Hoekstra, P., Oosterlaan, J., Heslenfeld, D., Buitelaar, J., & Mennes, M. (2016). Attention-Deficit/Hyperactivity Disorder symptoms coincide with altered striatal connectivity. Biological Psychiatry: Cognitive Neuroscience and Neuroimaging, 14, 353-363. https://doi.org/10.1016/J.BPSC.2016.03.008
Öngür, D., & Price, J. L. (2000). The organization of networks within the orbital and medial prefrontal cortex of rats, monkeys and humans. Cereb Cortex, 10(3), 206-219. https://doi.org/10.1093/cercor/10.3.206
Öngür, D., Ferry, A. T., & Price, J. L. (2003). Architectonic subdivision of the human orbital and medial prefrontal cortex. The Journal of Comparative Neurology, 460(3), 425–449. https://doi.org/10.1002/cne.10609
Padmanabhan, A., Lynch, C. J., Schaer, M., & Menon, V. (2017). The Default Mode Network in autism. Biological Psychiatry. Cognitive Neuroscience and Neuroimaging, 2(6), 476–486. https://doi.org/10.1016/j.bpsc.2017.04.004
Pikusa, M., & Jończyk, R. (2015). Functional abnormalities in Broca’s area in adolescents with ADHD: A resting-state fMRI study. Poznan Studies in Contemporary Linguistics, 51, 163 - 177. https://doi.org/10.1515/psicl-2015-0006
Pissiota, A., Frans, Ö., Fernández, M., Knorring, L., Fischer, H., & Fredrikson, M. (2000). Neurofunctional correlates of posttraumatic stress disorder: A PET symptom provocation study. European Archives of Psychiatry and Clinical Neuroscience, 252, 68-75. https://doi.org/10.1007/s004060200014
Price, J. L., & Drevets, W. C. (2010). Neurocircuitry of mood disorders. Neuropsychopharmacology: Official Publication of the American College of Neuropsychopharmacology, 35(1), 192–216. https://doi.org/10.1038/npp.2009.104
Radley, J. J., Arias, C. M., & Sawchenko, P. E. (2006). Regional differentiation of the medial prefrontal cortex in regulating adaptive responses to acute emotional stress. The Journal of Neuroscience, 26(50), 12967-12976. https://doi.org/10.1523/JNEUROSCI.4297-06.2006
Raichle, M. E., & Gusnard, D. A. (2002). Appraising the brain's energy budget. Proceedings of the National Academy of Sciences, 99(16), 10237-10239. https://doi.org/10.1073/pnas.172399499
Rajkowska, G., & Goldman-Rakic, P. S. (1995). Cytoarchitectonic definition of prefrontal areas in the normal human cortex: II. Variability in locations of areas 9 and 46 and relationship to the Talairach Coordinate System. Cerebral Cortex (1991), 5(4), 323–337. https://doi.org/10.1093/cercor/5.4.323
Rajmohan, V., & Mohandas, E. (2007). Mirror neuron system. Indian J Psychiatry, 49(1), 66-69. https://doi.org/10.4103/0019-5545.31522
Ramachandran, V. S. (2011). The tell-tale brain: A neuroscientist's quest for what makes us human. W. W. Norton & Company.
Rapanelli, M., Frick, L. R., & Zanutto, B. S. (2011). Learning an operant conditioning task differentially induces gliogenesis in the medial prefrontal cortex and neurogenesis in the hippocampus. PLoS ONE, 6(2), e14713. https://doi.org/10.1371/journal.pone.0014713
Rizzolatti, G., & Sinigaglia, C. (2008). Mirrors in the brain: How our mind share actions, emotions, and experience. Oxford University Press.
Sanei, S., & Chambers, J. A. (2013). EEG signal processing. John Wiley & Sons.
Sarnthein, J., Petsche, H., Rappelsberger, P., Shaw, G. L., & von Stein, A. (1998). Synchronization between prefrontal and posterior association cortex during human working memory. Proc Natl Acad Sci, 95(12), 7092-7096. https://doi.org/10.1073/pnas.95.12.7092
Sartory, G., Cwik, J., Knuppertz, H., Schürholt, B., Lebens, M., Seitz, R., & Schulze, R. (2013). In search of the trauma memory: A meta-analysis of functional neuroimaging studies of symptom provocation in Posttraumatic Stress Disorder (PTSD). PLoS ONE, 8. https://doi.org/10.1371/journal.pone.0058150
Scarr, E., Udawela, M., & Dean, B. (2019). Changed cortical risk gene expression in major depression and shared changes in cortical gene expression between major depression and bipolar disorders. Australian & New Zealand Journal of Psychiatry, 53, 1189 - 1198. https://doi.org/10.1177/0004867419857808
Schacter, D. L. (1977). EEG theta waves and psychological phenomena: A review and analysis. Biological Psychology, 5, 47-82. https://doi.org/10.1016/0301-0511(77)90028-x
Schoffelen, J. M., & Gross, J. (2009). Source connectivity analysis with MEG and EEG. Human Brain Mapping, 30(6), 1857-1865. https://doi.org/10.1002/hbm.20745
Schomer, D. L., & Lopes da Silva, F. H. (2011). Niedermeyer's electroencephalography: Basic principles, clinical applications, and related fields (6th ed.). Lippincott Williams & Wilkins.
Shang, J., Fu, Y., Ren, Z., Zhang, T., Du, M., Gong, Q., Lui, S., & Zhang, W. (2014). The Common Traits of the ACC and PFC in anxiety disorders in the DSM-5: Meta-analysis of voxel-based morphometry studies. PLoS ONE, 9. https://doi.org/10.1371/journal.pone.0093432
Sheline, Y. I., Price, J. L., Yan, Z., & Mintun, M. A. (2010). Resting-state functional MRI in depression unmasks increased connectivity between networks via the dorsal nexus. Proceedings of the National Academy of Sciences of the United States of America, 107(24), 11020–11025. https://doi.org/10.1073/pnas.1000446107
Silberstein, R., et al. (1993) unpublished data cited in Nunez, Neocortical dynamics and human EEG rhythms. Oxford University Press.
Sörös, P., Hoxhaj, E., Borel, P., Sadohara, C., Feige, B., Matthies, S., Müller, H., Bachmann, K., Schulze, M., & Philipsen, A. (2019). Hyperactivity/restlessness is associated with increased functional connectivity in adults with ADHD: A dimensional analysis of resting state fMRI. BMC Psychiatry, 19. https://doi.org/10.1186/s12888-019-2031-9
Stahl, S. M. (2008). Stahl’s essential psychopharmacology: Neuroscientific basis and practical applications (3rd ed.). Cambridge University Press.
Steriade, M. (2001). The intact and sliced brain. MIT.
Steriade, M. (2005). Cellular substrates of brain rhythms. In E. Niedermeyer, & F. Lopes da Silva (Eds.). Electroencephalography: Basic principles, clinical applications, and related fields (5th ed.). Lippincott Williams & Wilkins.
Steriade, M., & Llinás, R. R. (1988). The functional states of the thalamus and the associated neuronal interplay. Physiol Rev, 68(3), 649-742. https://doi.org/10.1152/physrev.1988.68.3.649
Sterman, M. B. (2000). EEG markers for attention deficit disorder: Pharmacological and neurofeedback applications. Child Study Journal, 30(1), 1-24.
Sterman, M. B., & Kaiser, D. A. (2001). Comodulation: A new QEEG analysis metric for assessment of structural and functional disorders of the CNS. Journal of Neurotherapy, 4(3), 73-83. https://doi.org/10.1300/J184v04n03_05
Stern, R. M., Ray, W. J., & Quigley, K. S. (2001). Psychophysiological recording (2nd ed.). Oxford University Press.
Streit, W. J. (2006). Microglial senescence: Does the brain's immune system have an expiration date? Trends in Neurosciences, 29(9), 506–510. https://doi.org/10.1016/j.tins.2006.07.001
Tarasi, L., & Romei, V. (2024). Individual alpha frequency contributes to the precision of human visual processing. Journal of Cognitive Neuroscience, 36(4), 602–613. https://doi.org/10.1162/jocn_a_02026
Thatcher, R. W. (2012). Coherence, phase differences, phase shift, and phase lock in EEG/ERP analyses. Developmental Neuropsychology, 37(6), 476-496. https://doi.org/10.1080/87565641.2011.619241
Thatcher, R. W., Biver, C. J., & North, D. M. (2009). EEG and brain connectivity: A tutorial. Unpublished manuscript.
Thompson, M., & Thompson, L. (2009). Asperger’s syndrome intervention: Combining neurofeedback, biofeedback, and metacognition. In T. H. Budzynski, H. K. Budzynski, J. R. Evans, & A. Abarbanel (Eds.). Introduction to quantitative EEG and neurofeedback (2nd ed.). Academic Press.
Thompson, M., & Thompson, L. (2015). The neurofeedback book: An introduction to basic concepts in applied psychophysiology (2nd ed.). Association for Applied Psychophysiology and Biofeedback.
Tiihonen, J., Kajola, M., & Hari, R. (1989). Magnetic mu rhythm in man. Neuroscience, 32(3), 793-800. https://doi.org/10.1016/0306-4522(89)90299-6
Traub, R. D., Miles, R., & Wong, R. K. S. (1989). Model of the origin of rhythmic population oscillations in the hippocampal slice. Science, 243, 1319-1325. https://doi.org/10.1126/science.2646715
van den Heuvel, M. P., & Sporns, O. (2011). Rich-club organization of the human connectome. J Neurosci, 31(44), 15775-15786. https://doi.org/10.1523/JNEUROSCI.3539-11.2011
Voloh, B., & Womelsdorf, T. (2016). A role of phase-resetting in coordinating large scale neural networks during attention and goal-directed behavior. Front Syst Neurosci, 10, https://doi.org/10.3389/fnsys.2016.00018
Voytek, B. (2013, May 20). Are there really as many neurons in the human brain as stars in the Milky Way? Brain Metrics. Scitable by Nature Education.
Warren, A. M., & McIlvane, W. J. (1998). Stimulus equivalence and the N400 effect. Poster presented at the 1998 Annual Meeting of the Cognitive Neuroscience Society in San Francisco, CA.
Wilson, J. (2003). Biological foundations of human behavior. Wadsworth/Thompson Learning.
Wilson, V. E., Thompson, M., Thompson, L., Thompson, J., Fallahpour, K., & Linden, M. K. (2011). Introduction to biofeedback (Neurofeedback). In B. W. Strack, M. K. Linden, & V. S. Wilson (Eds.). Biofeedback & neurofeedback applications in sport psychology. Association for Applied Psychophysiology and Biofeedback.
Winn, P. (Ed.). (2001). Dictionary of biological psychology. Routledge.
Xu, T., Yu, X., Perlik, A., Tobin, W., Zweig, J., Tennant, K., Jones, T., & Zuo, Y. (2009). Rapid formation and selective stabilization of synapses for enduring motor memories. Nature, 462(7275), 915-919.
Yang, Y., Ge, W., Chen, Y., Zhang, Z., Shen, W., Wu, C., Poo, M., & Duan, S. (2003). Contribution of astrocytes to hippocampal long-term potentiation through release of D-serine. Proceedings of the National Academy of Sciences of the United States of America, 100(25), 15194-15199. https://doi.org/10.1073/pnas.2431073100
Ye, T., Peng, J., Nie, B., Gao, J., Liu, J., Li, Y., Wang, G., Ma, X., Li, K., & Shan, B. (2012). Altered functional connectivity of the dorsolateral prefrontal cortex in first-episode patients with major depressive disorder. European Journal of Radiology, 81(12), 4035-4040. https://doi.org/10.1016/j.ejrad.2011.04.058
Zald, D. H., & Andreotti, C. (2010). Neuropsychological assessment of the orbital and ventromedial prefrontal cortex. Neuropsychologia, 48(12), 3377-3391. https://doi.org/10.1016/j.neuropsychologia.2010.08.012
Zhang, H., Watrous, A., Patel, A., & Jacobs, J. (2018). Theta and alpha oscillations are traveling waves in the human neocortex. Neuron. https://doi.org/10.1016/j.neuron.2018.05.019
Zhou, S., Su, S., Hong, A., Yang, C., Liu, Q., Feng, W., & Wang, Z. (2022). Abnormal functional connectivity of brain regions associated with fear network model in panic disorder. The World Journal of Biological Psychiatry, 23, 764 - 772. https://doi.org/10.1080/15622975.2022.2038389
Zhou, Y., Liang, M., Jiang, T., Tian, L., Liu, Y., Liu, Z., Liu, H., & Kuang, F. (2007). Functional dysconnectivity of the dorsolateral prefrontal cortex in first-episode schizophrenia using resting-state fMRI. Neuroscience Letters, 417, 297-302. https://doi.org/10.1016/j.neulet.2007.02.081
Zhukovsky, P., Anderson, J., Coughlan, G., Mulsant, B., Cipriani, A., & Voineskos, A. (2020). Coordinate-based network mapping of brain structure in Major Depressive Disorder in younger and older adults: A systematic review and meta-analysis. The American Journal of Psychiatry, appiajp202121010088. https://doi.org/10.2139/ssrn.3730007