Neurophysiology
What You Will Learn in This Chapter
This chapter explores the electrical and chemical foundations of the brain that make biofeedback and neurofeedback possible. You will discover how neurons communicate through action potentials and synaptic transmission, learn the supporting roles of glial cells, and understand how the electroencephalograph (EEG) captures the brain's electrical activity from the scalp.
Along the way, you will examine event-related potentials, slow cortical potentials, and the neuroplasticity that underpins neurofeedback training. By the end of this chapter, you will be able to explain how local field potentials generate the EEG signal, why the EEG reflects postsynaptic potentials rather than action potentials, and how long-term depression and long-term potentiation reshape neural circuits during learning.
BCIA Blueprint Coverage: This unit addresses II. Basic Neurophysiology and Neuroanatomy, A. Neurophysiology. It covers the Bioelectric Origin and Functional Correlates of EEG, Definition of ERPs and SCPs, and Neuroplasticity.
The brain is the hardest-working organ in the body. A typical brain weighs about 3 pounds (Chan et al., 2009) and consists of about 2% of body weight, yet it consumes 20% of the body's energy and oxygen (Raichle & Gusnard, 2002). This extraordinary metabolic demand produces electrical fields that are measurable from the scalp and form the basis of neurofeedback. Child graphic © vasara/Shutterstock.com.
The brain accomplishes its work through a sophisticated communication and command-and-control system that monitors and manages interactions between roughly 86 billion neurons, with cortical neurons carrying roughly 5,000-10,000 synaptic connections each and total synapse counts for the adult brain usually placed on the order of 100 trillion (Breedlove & Watson, 2023). For clinicians, this immense connectivity means that even small changes in neural communication patterns can produce clinically meaningful effects.
An electroencephalograph (EEG) monitors brainwave activity at various frequencies, from DC shifts (slow cortical potentials) to fast potentials exceeding 50 Hz. The EEG records the excitatory postsynaptic potentials (EPSPs) and inhibitory postsynaptic potentials (IPSPs) propagated by the apical dendrites of large pyramidal cells arranged in thousands of cortical columns. Local field potentials, the aggregate effect of interconnected neuron firing and modulation by glial cells, regulate neuron excitability and firing. Action potential animation © NIMEDIA/Shutterstock.com.
Neuroplasticity, the remodeling of neurons and neural networks with experience, is responsible for learning and memory and makes neurofeedback training possible. Without neuroplasticity, operant conditioning of brainwave activity would have no lasting effect.
This unit addresses II. Basic Neurophysiology and Neuroanatomy - A. Neurophysiology.

This unit covers the Bioelectric Origin and Functional Correlates of EEG, Definition of ERPs and SCPs, and Neuroplasticity.
🎧 Chapter Lecture: Neurophysiology
Key Concept
The scalp EEG does not record the action potentials that neurons fire. It reflects the summed excitatory and inhibitory postsynaptic potentials (EPSPs and IPSPs) generated in the apical dendrites of large pyramidal cells, which are arranged in thousands of cortical columns and polarized synchronously. Because these slower postsynaptic potentials, rather than the brief action potentials, reach the scalp, everything a clinician trains during neurofeedback is a downstream reflection of synaptic communication. Keeping this distinction in mind explains why this chapter devotes so much attention to synapses, dendrites, and local field potentials.
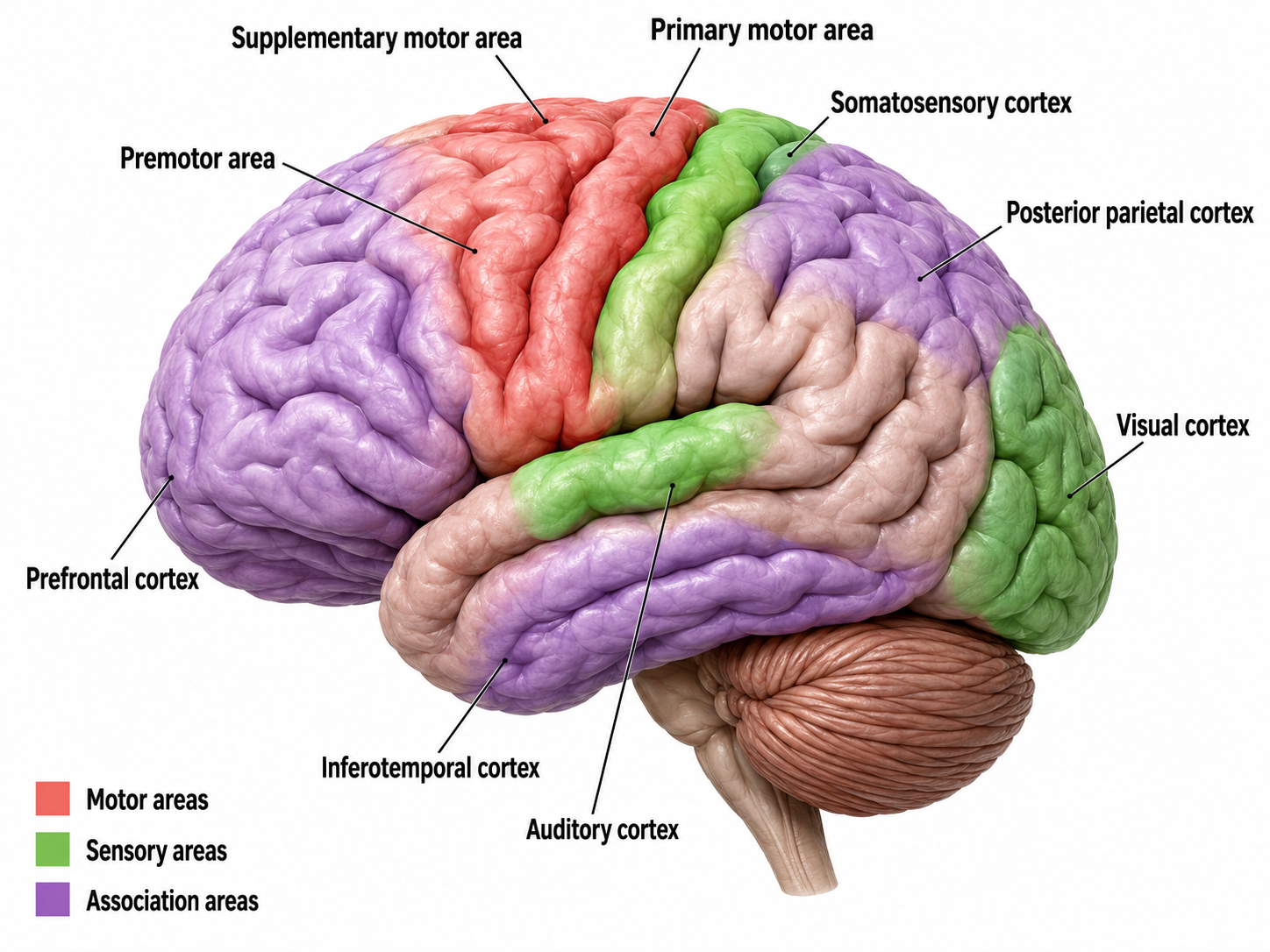
Bioelectric Origin and Functional Correlates of EEG
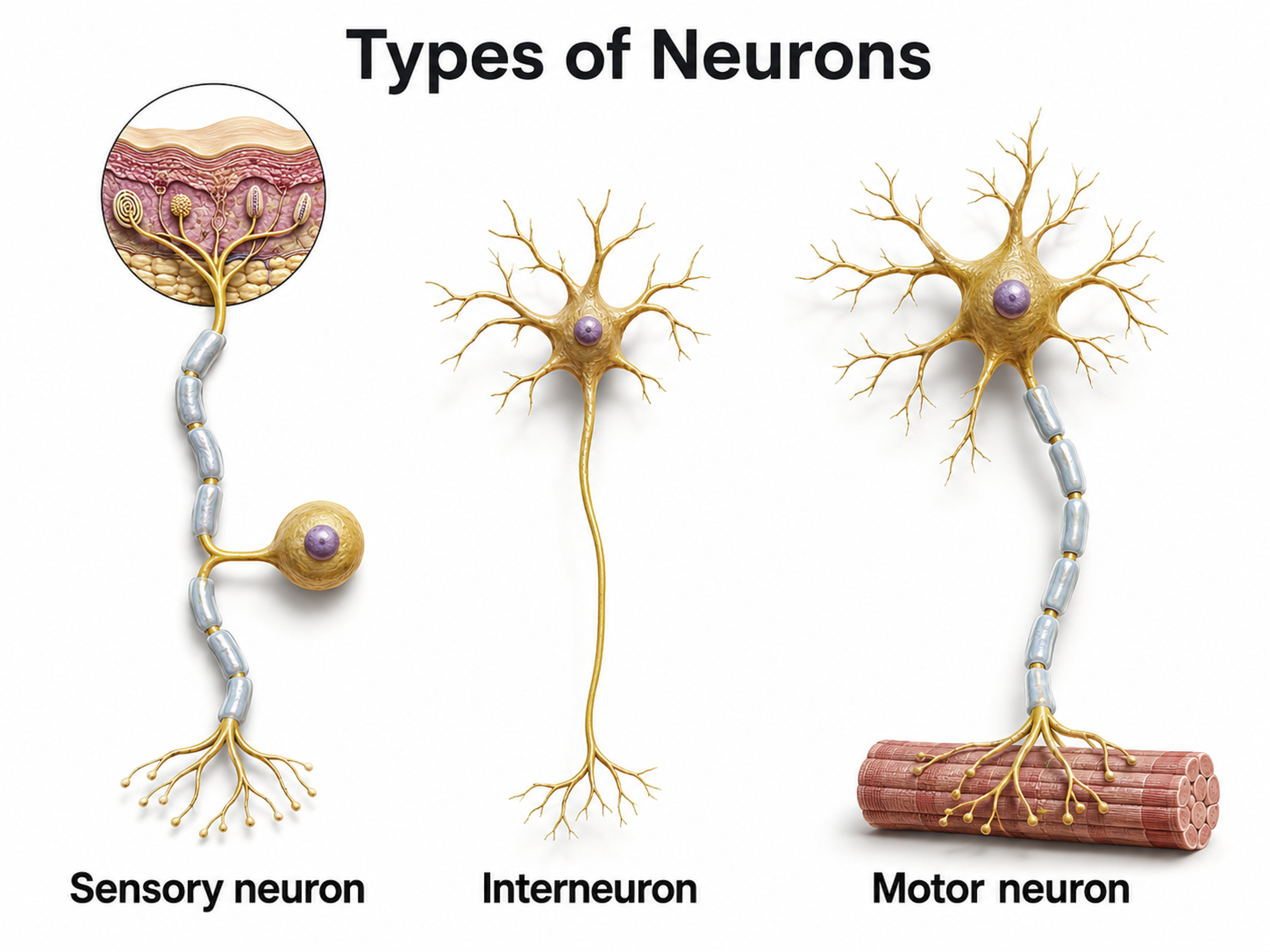
Types of Neurons
This section covers the three major types of neurons, sensory, motor, and interneurons, along with the glial cells that work in partnership with them. Understanding these cell types is essential because each plays a distinct role in generating the electrical signals that neurofeedback clinicians measure and train. Neuron graphic © SciePro/Shutterstock.com.
We can divide neurons into three functional categories: sensory, motor, and interneurons. Glial cells like astrocytes and microglia work in partnership with neurons to support communication and maintain the neural environment.
Sensory neurons are specialized for receiving information from the environment and the body. They are called afferent because they transmit sensory information toward the central nervous system (brain and spinal cord). Whether a client feels the warmth of a thermal biofeedback sensor or hears an auditory neurofeedback tone, sensory neurons relay that information centrally for processing. Sensory neuron graphic © TimeLineArtist/Shutterstock.com.
Motor neurons convey commands to glands, muscles, and other neurons. They are called efferent because they carry information toward the periphery. When a client consciously relaxes the frontalis muscle during surface EMG biofeedback, motor neurons translate that intention into reduced muscle tension. Motor and sensory neuron graphic © iso-form llc/Shutterstock.com.
.jpg)
Interneurons provide the integration required for decisions, learning and memory, perception, planning, and movement. They have short processes, analyze incoming information, and distribute their analysis within neural networks. Interneurons are largely confined to the central nervous system, though they also occur in the enteric nervous system and in peripheral autonomic ganglia. Under the broad definition, any neuron that is neither sensory nor motor, they account for the great majority of neurons (Breedlove & Watson, 2023); in the neocortex, local-circuit interneurons make up roughly 20-30% of neurons, the remainder being pyramidal projection cells.
Local interneurons analyze small amounts of information provided by neighboring neurons. Relay interneurons connect networks of local interneurons from separate brain regions, enabling diverse functions like perception, learning, memory, and executive functions such as planning (Carlson & Birkett, 2021). This local-to-global architecture helps explain why neurofeedback training at a single electrode site can influence complex cognitive and emotional processes. Neuron graphic adapted from © Aldona Griskeviciene/Shutterstock.com.
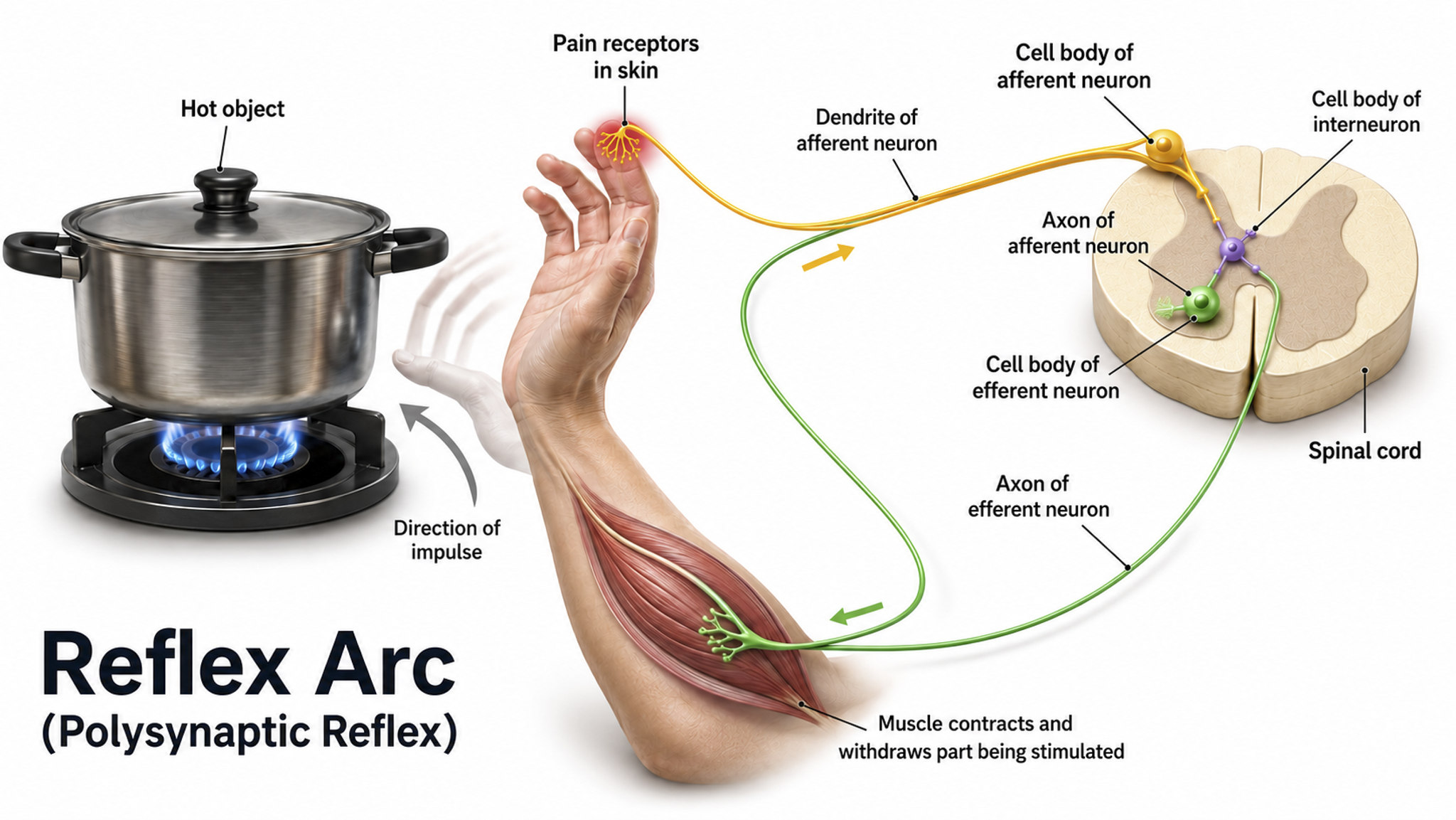
Reflex arc graphic adapted from © SANDIP NEOGI/Shutterstock.com.
Neuron Structure
This section examines the key structural components of neurons, from the cell body to terminal buttons. These structures work together to receive, integrate, and transmit electrical and chemical signals. Understanding neuron anatomy is fundamental to grasping how the EEG signal originates and what neurofeedback clinicians are ultimately measuring.
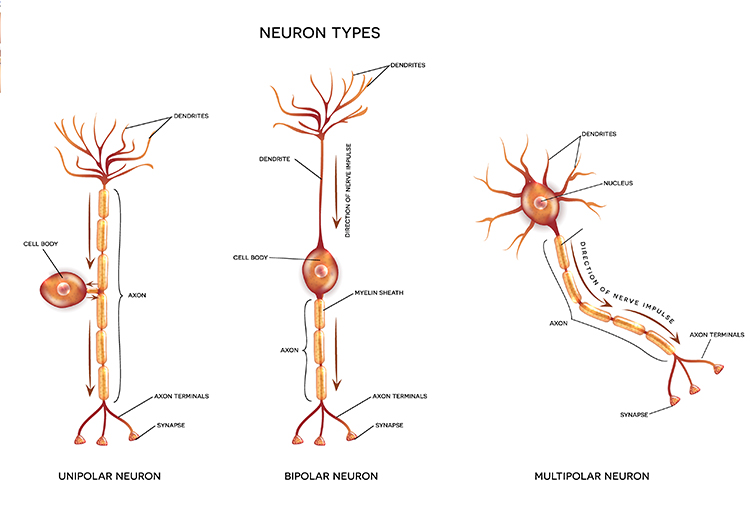
While neurons have over 200 different designs to perform specialized jobs in the nervous system, they generally share five structures: a cell body or soma, dendrites, an axon hillock, an axon, and terminal buttons.
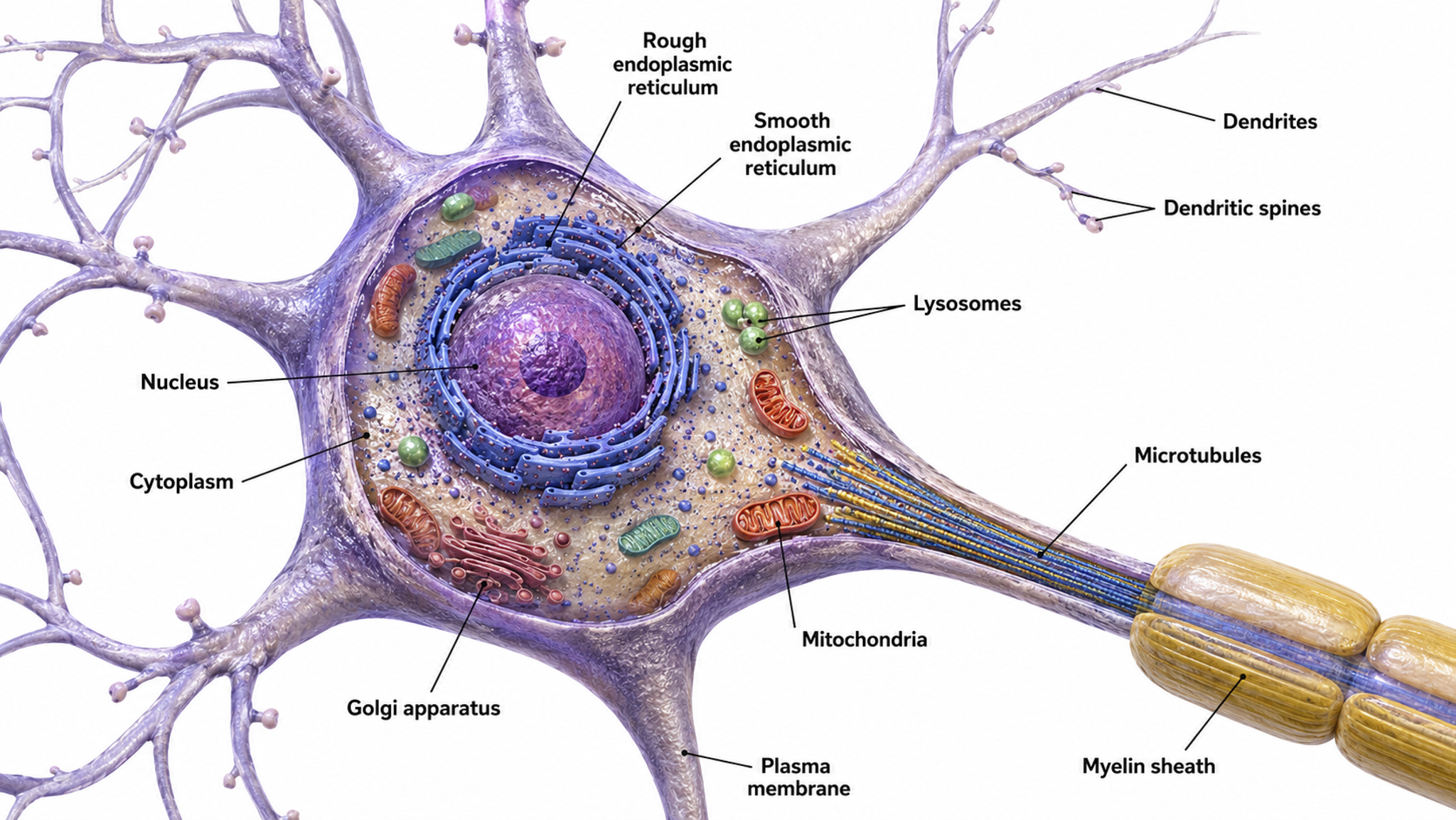
The cell body or soma contains the machinery for the neuron's life processes. It receives and integrates EPSPs and IPSPs, small graded positive and negative changes in membrane potential generated by axons. The cell body of a typical neuron is 20 μm in diameter, and its spherical nucleus, which contains chromosomes comprised of DNA, is 5-10 μm across.
The cell body is the primary location where neurons manufacture proteins (like enzymes, receptors, and ion channels) and peptides (neurotransmitters like oxytocin), though there is increasing evidence of distributed protein manufacturing via local mRNA translation (Nagano & Araki, 2021). Check out the Khan Academy YouTube video, Anatomy of a Neuron. Cell body graphic © MattL_Images/Shutterstock.com.
Mitochondria power diverse processes throughout neurons, including opening ion channels, conducting action potentials, releasing and returning neurotransmitters, and transporting proteins. They are directly responsible for EEG signal strength since they fuel the postsynaptic potentials that scalp electrodes detect. Mitochondria are orange in the graphic below © Corona Borealis Studio/Shutterstock.com.
Mitochondria are far more than simple cellular energy factories. In the brain, these organelles are master multitaskers whose involvement in cellular respiration, calcium homeostasis, and reactive oxygen species (ROS) management makes them integral to the brain's demanding physiological environment (Fischer et al., 2020). At their most fundamental level, mitochondria generate the ATP that neurons desperately need. Neurons are constantly sending electrical signals, releasing neurotransmitters, and rebuilding connections, which is why mitochondria cluster at synapses like miniature power plants, ensuring there is always enough fuel for crucial processes like learning and memory formation (Harris et al., 2012).
Mitochondria also serve as cellular calcium regulators, maintaining strict control over calcium levels inside neurons. This calcium management is critical for proper neural signaling; when mitochondria fail at this job, it can trigger a cascade of problems that may lead to neurodegenerative diseases. During intense neural activity, mitochondria buffer incoming calcium waves, preventing toxic buildups while fine-tuning the cellular responses that drive neurotransmitter release and gene expression (Brini et al., 2014). These organelles also walk a delicate tightrope with reactive oxygen species (ROS), which they generate as a byproduct of energy production. In small amounts, ROS serve as important signaling molecules, but excessive production can damage proteins, lipids, and DNA, a particular danger in the brain, which is extremely vulnerable to oxidative damage due to its high oxygen consumption and abundance of lipids (Lin & Beal, 2006).
Mitochondria are also cellular life-and-death decision makers, playing a crucial role in apoptosis (programmed cell death). This process is essential during brain development, helping to sculpt neural circuits by eliminating unnecessary neurons through specific pathways involving cytochrome c release and caspase activation (Green et al., 2011). When damaged, mitochondria can release danger signals called DAMPs (damage-associated molecular patterns), including mitochondrial DNA, which can trigger inflammatory responses that contribute to conditions like multiple sclerosis and chronic traumatic encephalopathy (West et al., 2015). As we age, mitochondrial function gradually declines, leading to decreased ATP production and increased ROS generation, which affects crucial processes like synaptic function and memory formation (Lopez-Otin et al., 2013).
The dynamic nature of mitochondria is particularly fascinating in the context of brain plasticity. These organelles constantly undergo fusion and fission, processes that allow them to adapt to changing energy demands and repair damage. These dynamics are especially important in maintaining synaptic plasticity, the foundation of learning and memory that makes neurofeedback training possible. When these processes malfunction, it can lead to developmental disorders including autism spectrum disorder and intellectual disabilities (López-Doménech et al., 2016). For neurofeedback clinicians, understanding mitochondrial function helps explain why factors like sleep quality, exercise, and nutrition can profoundly influence a client's capacity for neural learning during training.
Dendrites are branched structures designed to receive messages from other neurons via axodendritic synapses (junctions between axons and dendrites) and send messages to other neurons via dendrodendritic synapses (junctions between the dendrites of two neurons). Dendrites receive thousands of synaptic contacts and have specialized proteins called receptors for neurotransmitters released into the synaptic cleft (Bear, Connors, & Paradiso, 2026).
A neuron's dendrites are called a dendritic tree, and each extension is called a dendritic branch.
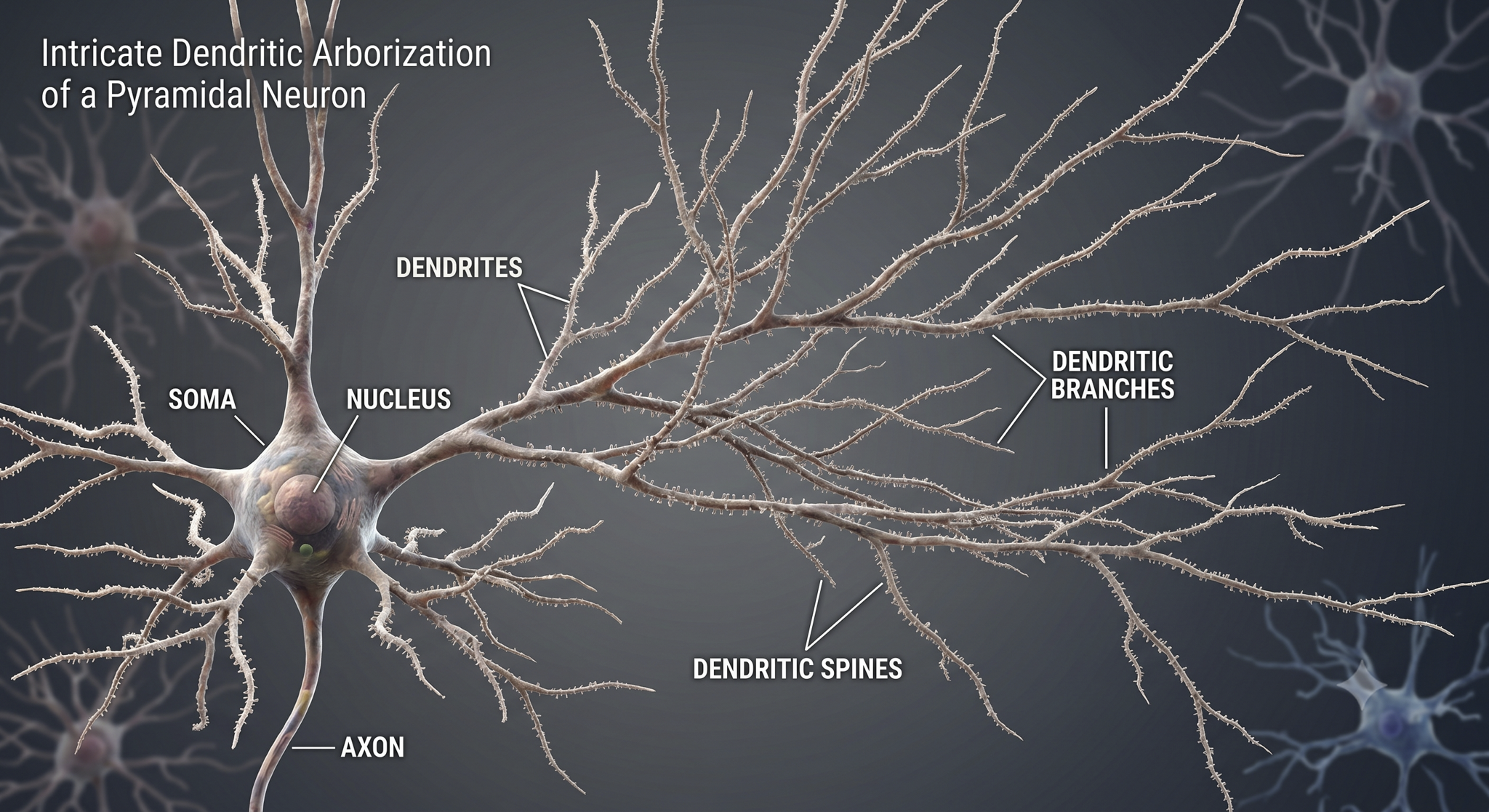
Biological psychologists classify neurons based on whether their dendrites feature spines. Dendritic spines are protrusions on the dendrite shaft where axons typically form axodendritic synapses. These tiny structures are critically important in neurofeedback because they are a primary site of synaptic plasticity; they can grow, shrink, or change shape in response to experience, including neurofeedback training. Graphic © Jose Luis Calvo/Shutterstock.com.
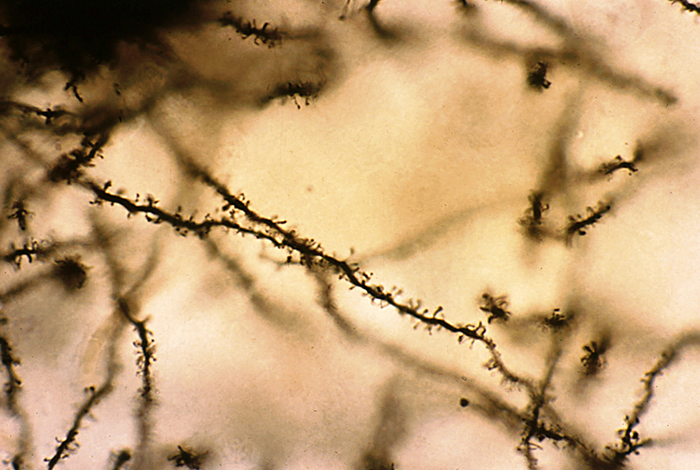
Spiny neurons have dendritic spines, while aspinous neurons do not (Bear, Connors, & Paradiso, 2026).
During learning, spines' number, size, and shape may change to adjust the space for receptors (neuroplasticity).
An axon is a cylindrical structure only found in neurons that is specialized for distributing information within the central and peripheral nervous systems. Axons range from 1 to 25 µm in diameter and 0.1 mm to more than a meter in length. Most neurons are interneurons, and many have axons and dendrites short enough that they do not extend beyond their local cell cluster. Axons usually branch repeatedly, and each branch is called an axon collateral.
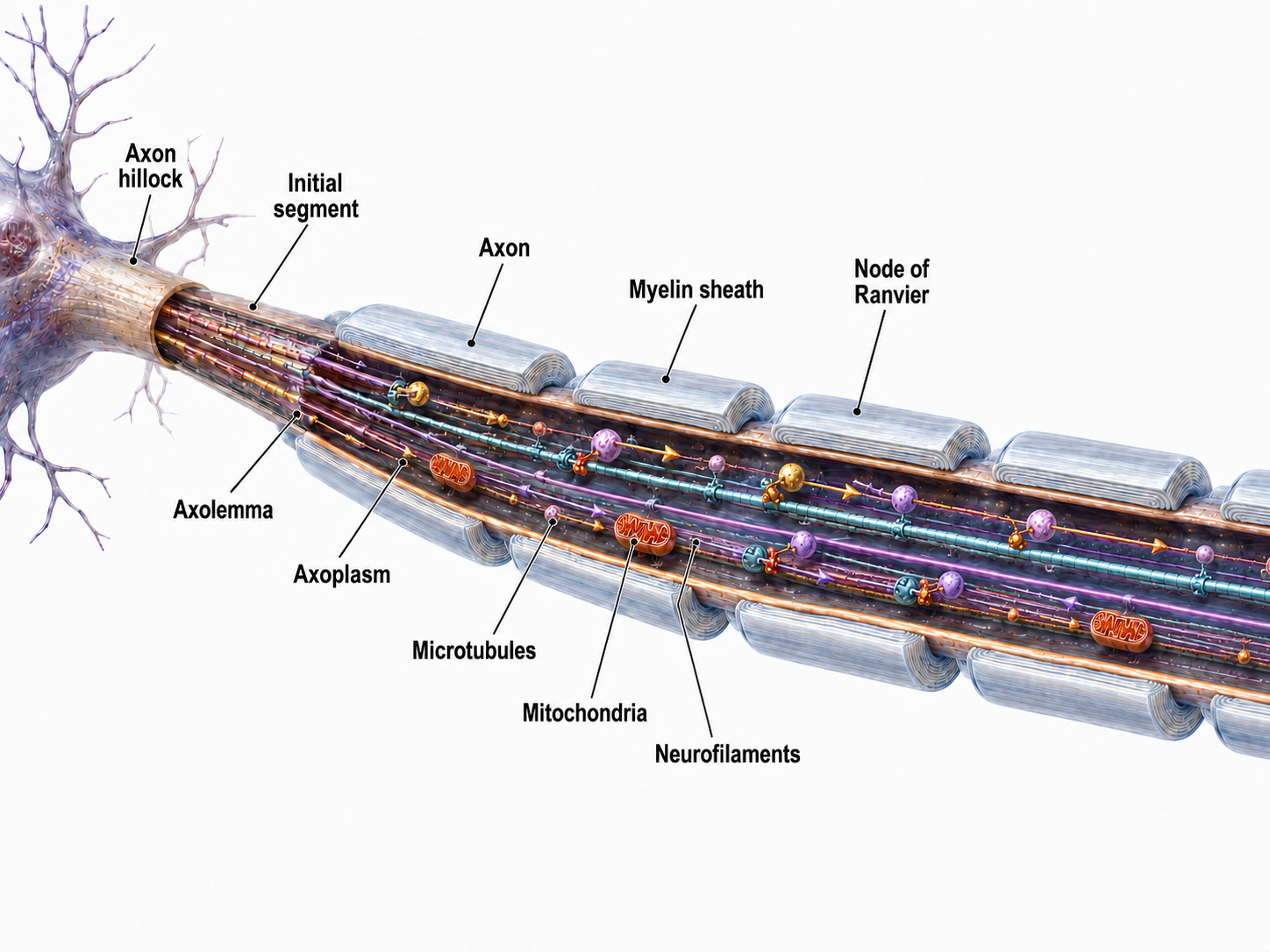
Axons transmit action potentials toward a neuron's terminal buttons. Using microtubules, an axon also bidirectionally transports molecules between the cell body and terminal buttons.
An axon hillock is a swelling of the cell body where the axon begins. Think of it as the neuron's decision point; it integrates all incoming signals to determine whether to fire. The middle of an axon is the axon proper, and the end is the axon terminal (Bear, Connors, & Paradiso, 2026).
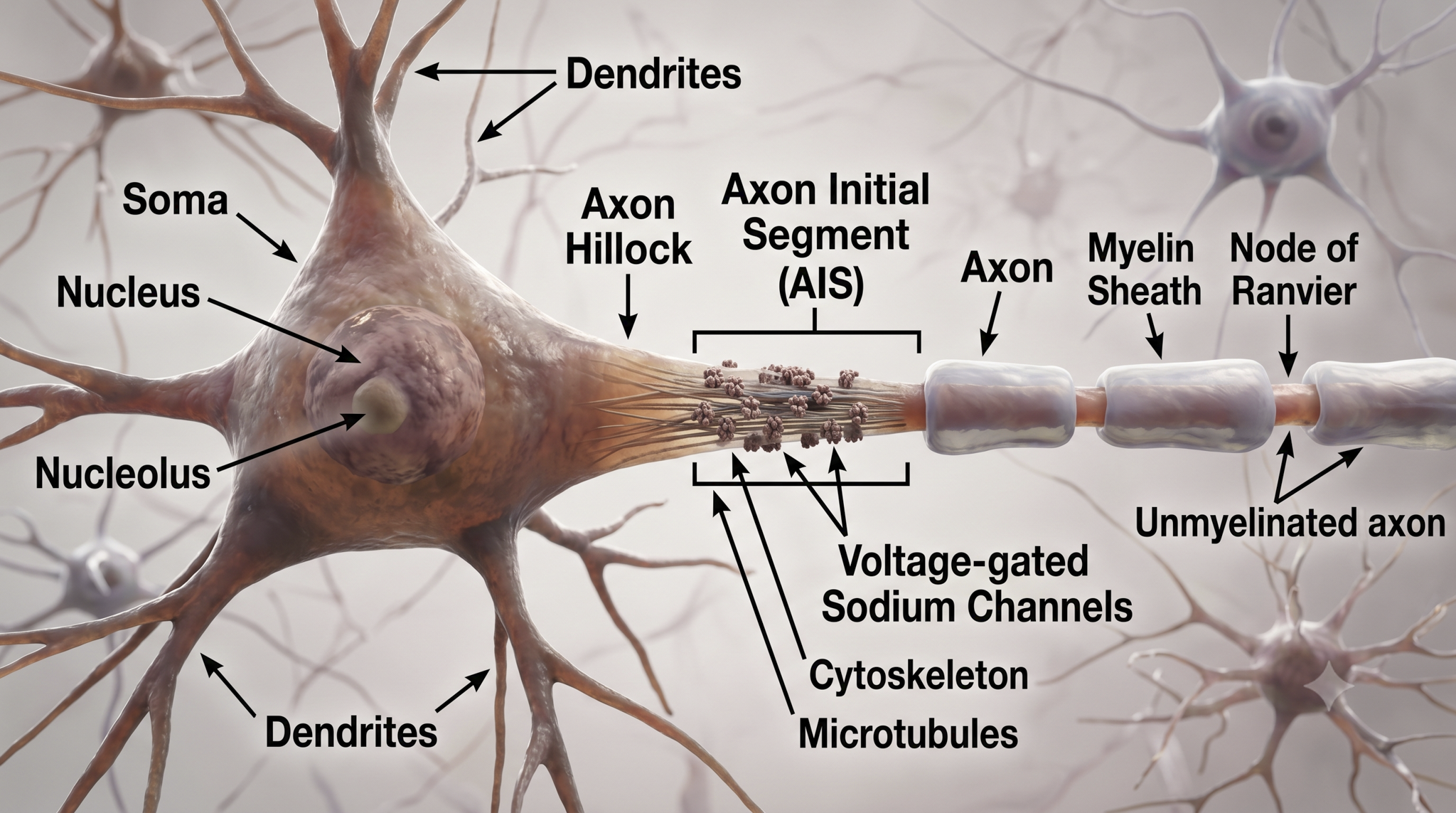
The axon hillock sums EPSPs and IPSPs over milliseconds to generate an action potential.
Axon terminals are buds located on the ends of axon branches that form synapses and release neurochemicals to other neurons. Axon terminals contain vesicles that store neurotransmitters for release when an action potential arrives. Their presynaptic membrane may have reuptake transporters that return neurotransmitters (NTs) from the synapse or extracellular space for repackaging.
Types of Glial Cells
This section covers the four main categories of glial cells: astrocytes, microglia, oligodendrocytes, and Schwann cells. Once dismissed as mere scaffolding, glial cells are now recognized as active partners in neural processing. Their role in modulating neuron excitability is directly relevant to understanding EEG generation and the mechanisms behind neurofeedback.
While there are hundreds of types of neurons, there are only four main categories of glial cells (astrocytes, microglia, oligodendrocytes, and Schwann cells).
Old school view: glial cells mainly provide structural support (glia is derived from the Greek for glue).
New school view: glial cells help neurons process information, including modulating neuron excitability.
Check out the YouTube video, Neurology - Glial Cells, White Matter and Gray Matter.
Astrocytes are star-shaped glial cells in the central nervous system that perform vital functions reaching well beyond structural support. Astrocyte endfeet form junctions with capillaries comprising part of the protective blood-brain barrier, and they regulate blood flow to neurons, delivering stored glucose during peak metabolic demand. Schummers et al. (2008) showed that cortical astrocytes carry tuned sensory responses and shape the hemodynamic signal, one route by which they take part in this coupling. Astrocyte graphic © Kateryna Kon/Shutterstock.com.
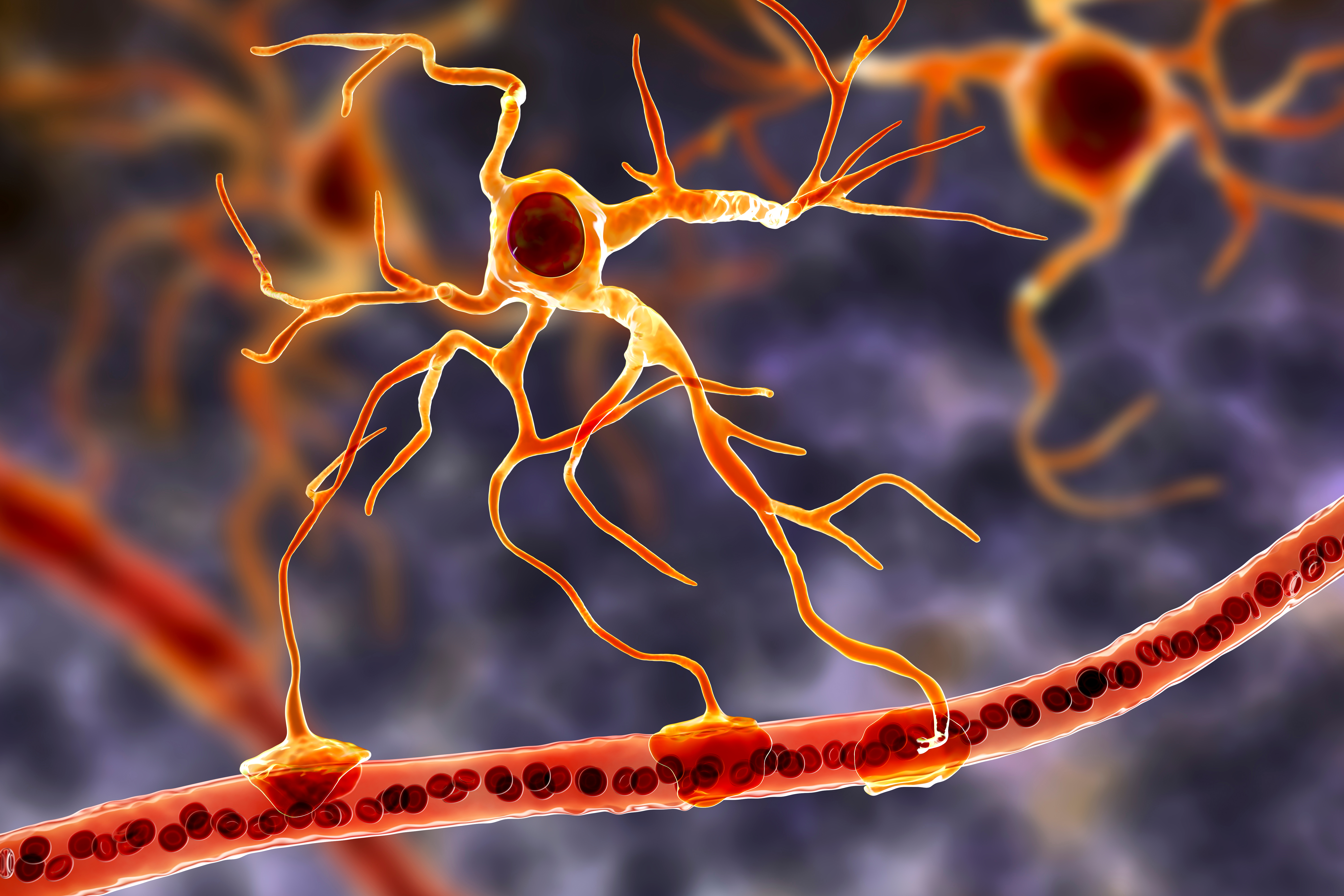
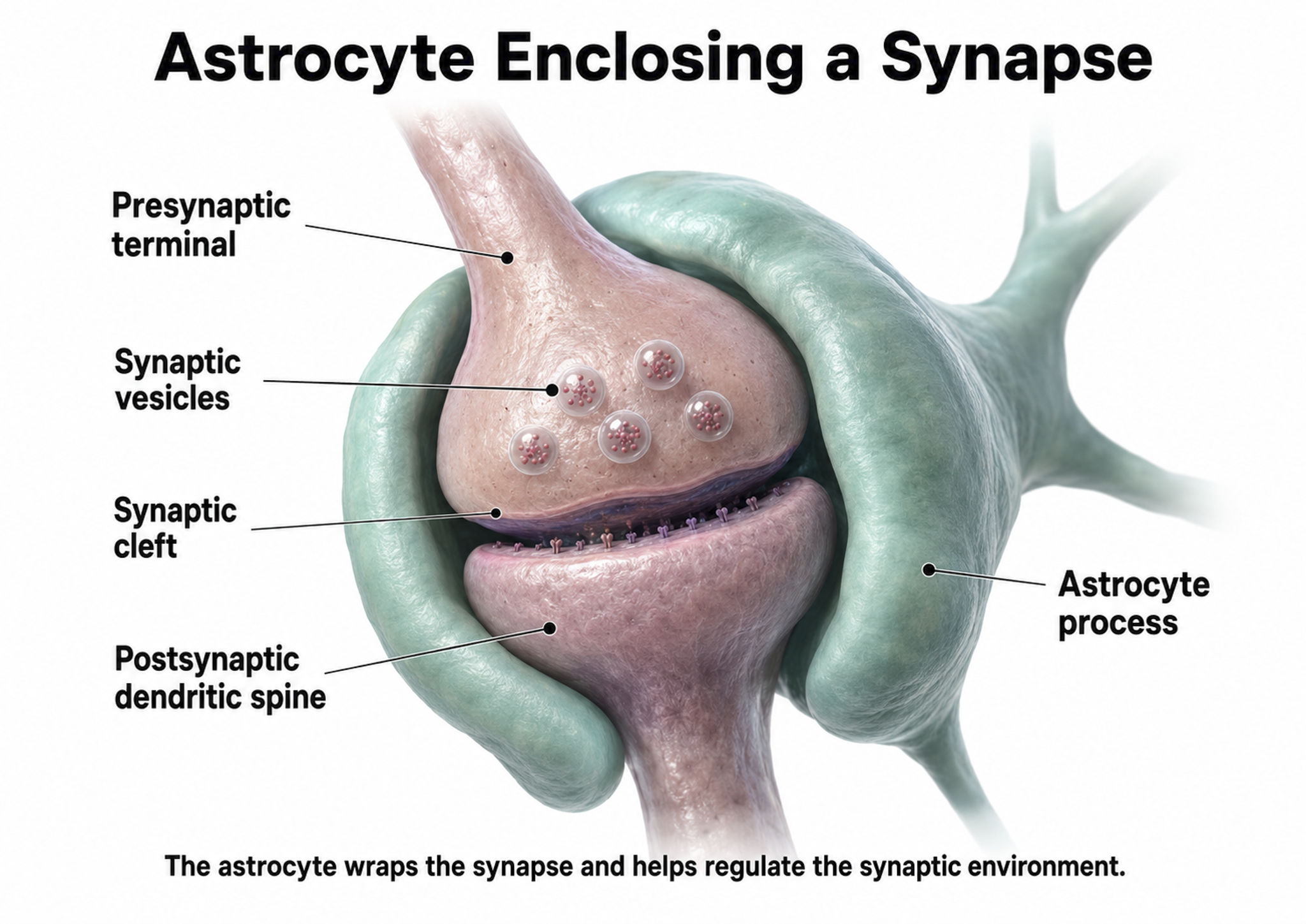
Astrocytes enclose synapses, determine where synapses can form by releasing specialized molecules, regulate synapse maturation, bidirectionally communicate with synapses, prune surplus synapses, help neurons regulate brain microcirculation, and eavesdrop on nearby synapse activity (Breedlove & Watson, 2023; Parri & Crunelli, 2003; Shan et al., 2021). Astrocytes also transport amino acid NTs (e.g., GABA and glutamate) from the synaptic cleft.
Astrocytes are theorized to participate in gliotransmission between neurons and each other (Eroglu & Barres, 2010; Perea et al., 2009). However, gliotransmission remains controversial.
. . . the physiological role of gliotransmission is highly debatable . . . as gliotransmitter release has been reliably demonstrated only in vitro in cultures and brain slice experiments that are often accompanied by manipulations (e.g., high frequency stimulation) which can affect astrocytic channels or receptors leading to impaired signaling cascades. This experimental design imposes questions about the existence of gliotransmission . . . and whether it plays a physiological role in the brain . . . (Buskila et al., 2019).
The presynaptic and postsynaptic neurons and astrocytes comprise a tripartite synapse, a three-part communication unit that expands our understanding of how neural signals are generated and modified. This concept is important for neurofeedback because it means the EEG signal reflects not just neuron-to-neuron communication, but a more complex interaction involving glial modulation.
An essential role of astrocytes is regulating the chemical content of this extracellular space. For example, astrocytes envelop synaptic junctions in the brain, thereby restricting the spread of neurotransmitter molecules that have been released. Astrocytes also have special proteins in their membranes that actively remove many neurotransmitters from the synaptic cleft. A recent and unexpected discovery is that astrocytic membranes also possess neurotransmitter receptors that, like the receptors on neurons, can trigger electrical and biochemical events inside the glial cell (Bear et al., 2020, p. 49).
![]()
Astrocyte glutamate release may be essential for hippocampal long-term depression (LTD), a long-lasting reduction in transmission strength, and long-term memory modulation (Navarrete et al., 2019). Astrocyte calcium and brain-derived neurotrophic factor (BDNF) release appear critical for late-phase hippocampal long-term potentiation (LTP), a long-lasting increase in transmission strength, and long-term memory regulation (Liu et al., 2021).
Astrocytes communicate with each other through gap junctions (Bennett et al., 2003). They may also contribute to brainwaves by regulating synapses via these gap junctions and calcium signaling.
These capabilities allow astrocytes to regulate neuronal excitability via glutamate uptake, gliotransmission and tight control of the extracellular positively-charged potassium (K+) levels via a process termed K+ clearance. Spatio-temporal synchrony of activity across neuronal and astrocytic networks, both locally and distributed across cortical regions, underpins brain states and thereby behavioral states, and it is becoming apparent that astrocytes play an important role in the development and maintenance of neural activity underlying these complex behavioral states (Buskila et al., 2019).
Microglia
Microscopic microglial cells participate in the immune response and are the brain's resident defense force. They scavenge and engulf diverse materials (phagocytosis), release cytotoxins to control infection, present antigens to T-cells, remove branches from neurons near damaged tissue to aid regrowth (synaptic stripping), promote tissue repair, and if they are persistently activated, they can promote chronic neuroinflammation in the CNS that amplifies neurodegeneration. They assist synaptic remodeling by removing unnecessary synapses. Finally, microglia cross the blood-brain barrier to promote homeostasis (Bear, Connors, & Paradiso, 2026). Graphic © Juan Gaertner/Shutterstock.com.
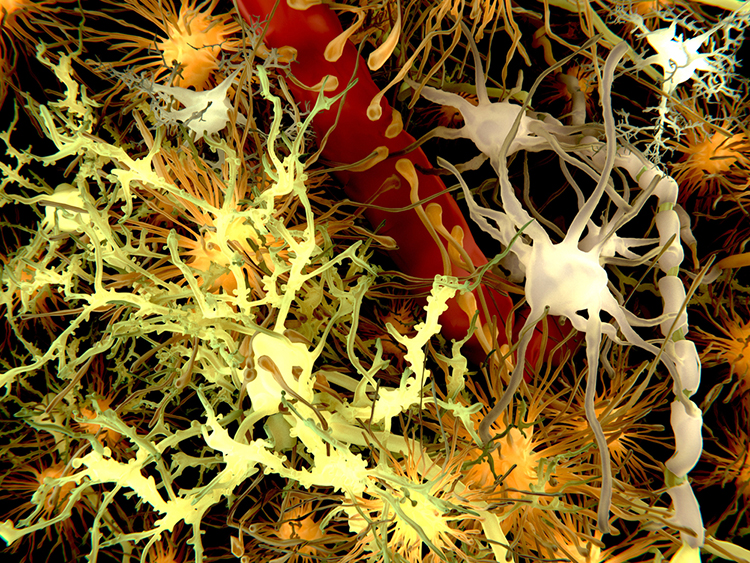
Description: yellow = neurons, orange = astrocytes, grey = oligodendrocytes, white = microglia.
Oligodendrocytes, which are smaller than astrocytes, form up to 50 segments of myelin that only insulate adjacent axons within the brain and spinal cord of the central nervous system.
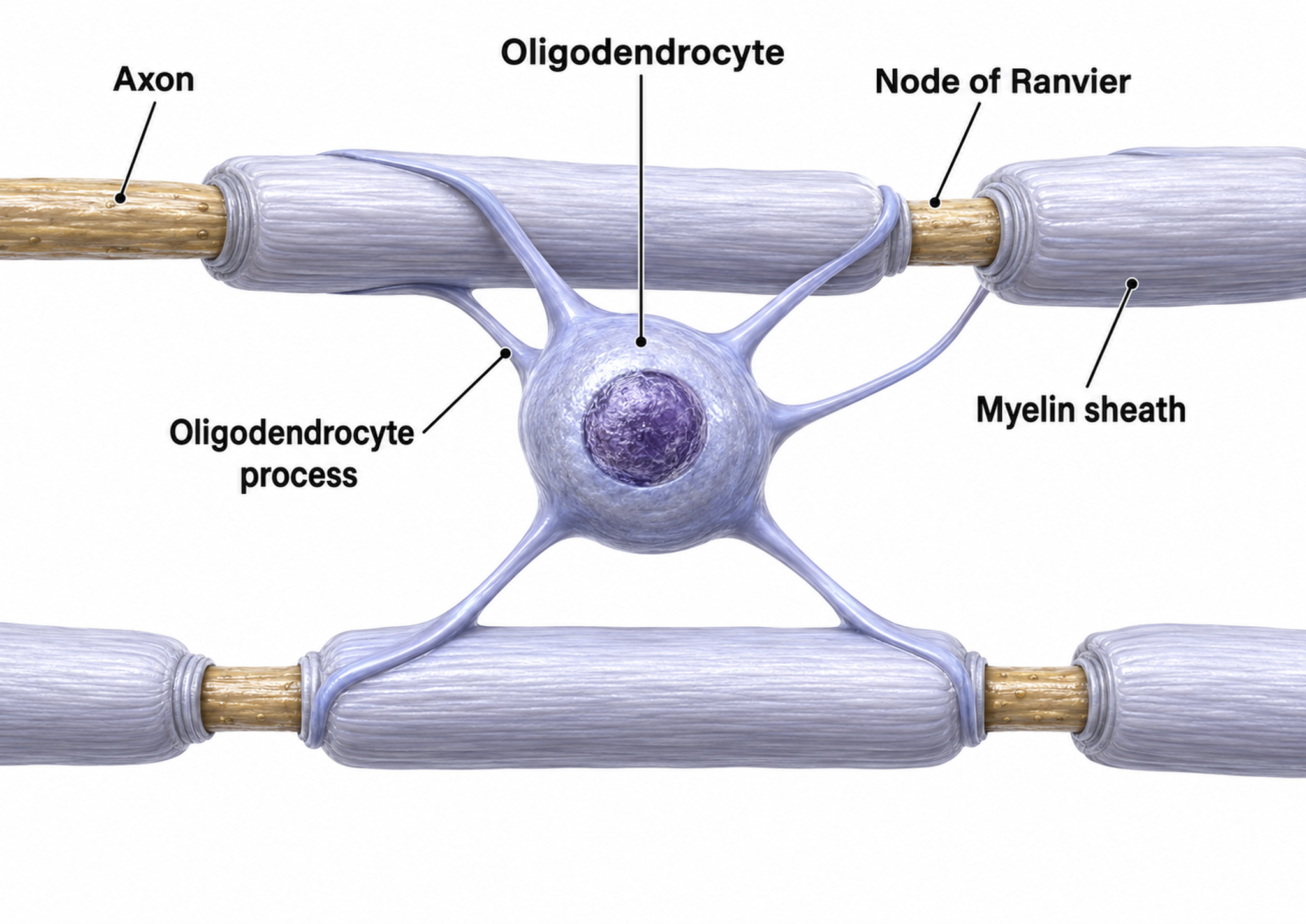
Oligodendrocytes block axonal regeneration by releasing growth inhibitory proteins, which partly explains the minimal functional recovery in the CNS following spinal cord damage.
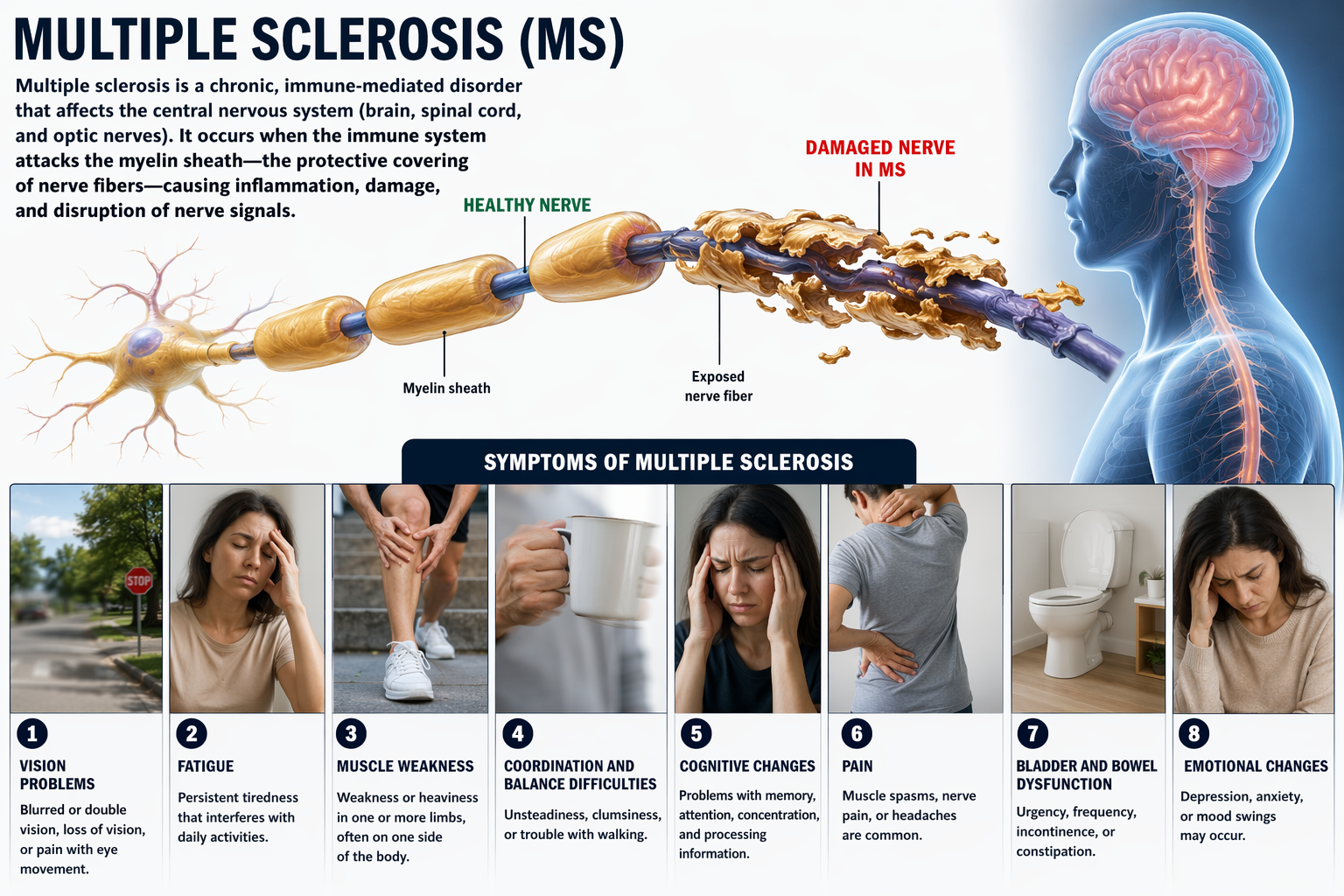
Multiple sclerosis, a demyelinating disease, destroys oligodendrocytes, disrupting neural communication in ways that clinicians can sometimes detect through QEEG assessment.
Schwann cells provide myelin for single peripheral nervous system (PNS) axons and facilitate axonal regeneration following damage (Breedlove & Watson, 2023).
The Resting Membrane Potential
This section explains the resting membrane potential, the steady electrical charge a neuron maintains when it is not signaling. Understanding the resting potential is foundational because every postsynaptic potential, action potential, and ultimately the EEG itself is a deviation from this baseline. When clinicians watch a neurofeedback display, they are watching the summed consequences of millions of neurons departing from and returning to this resting state.
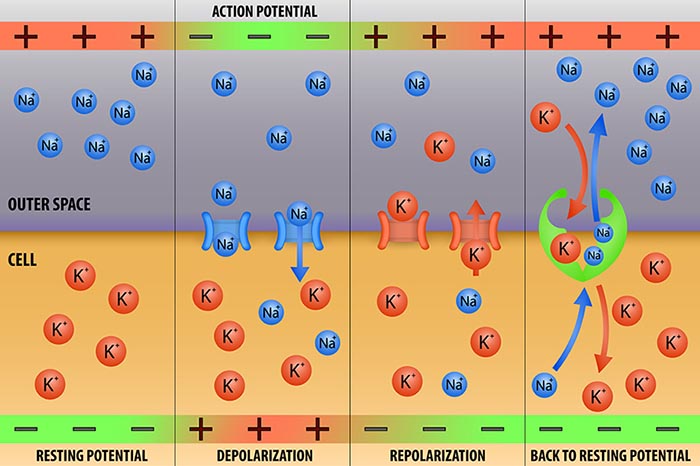
The resting membrane potential is the voltage difference across a neuron's membrane at rest, typically about -70 mV, with the inside of the cell more negative than the outside. This charge separation exists because ions are distributed unequally across the membrane. Sodium (Na+) and chloride (Cl-) ions are concentrated outside the neuron, while potassium (K+) ions and large negatively charged proteins are concentrated inside (Bear, Connors, & Paradiso, 2026). This uneven distribution is the ion concentration gradient that stores the energy driving neural signaling.
Two forces hold the resting potential in place: selective permeability and active transport. Selective permeability means the resting membrane is far more permeable to potassium than to sodium because potassium leak channels stay open. Potassium therefore drifts out of the cell down its concentration gradient, leaving the interior more negative until the growing electrical attraction pulling potassium back inward balances the chemical push driving it outward. The membrane voltage at which these two forces cancel for a given ion is its equilibrium potential, and because the resting membrane is dominated by potassium, the resting potential sits close to potassium's equilibrium value (Breedlove & Watson, 2023).
Left alone, the small constant leak of ions would gradually erase these gradients. The sodium-potassium pump prevents this by using ATP to exchange three sodium ions pushed out of the cell for every two potassium ions brought in, continuously restoring the gradients that the resting potential depends on. This relentless transport is one reason neurons are such heavy energy consumers and why the mitochondria described earlier cluster where this demand is greatest (Breedlove & Watson, 2023).
Clinical Application
Because the resting potential is actively maintained, anything that limits a neuron's energy supply degrades it. Hypoxia, ischemia, and severe hypoglycemia starve the sodium-potassium pump, the gradients run down, and cortical signaling slows. This is the physiological reason such states produce diffuse slowing in the EEG, a pattern clinicians may encounter when a client's medical status, medication, or substance use compromises cortical metabolism.
Excitatory and Inhibitory Postsynaptic Potentials
This section explains the small graded potentials, EPSPs and IPSPs, that are the primary source of the EEG signal recorded at the scalp. Understanding these potentials is essential because they, not action potentials, are what neurofeedback clinicians measure during neurofeedback sessions.
Graded positive and negative changes in membrane potential, called excitatory postsynaptic potentials and inhibitory postsynaptic potentials, are essential to the EEG and communication among neurons.
An excitatory postsynaptic potential (EPSP) is a subthreshold depolarization that makes the membrane potential more positive and pushes the neuron toward its excitation threshold. EPSPs are produced when neurotransmitters bind to receptors and cause positive sodium ions to enter the cell. At a single synapse, a postsynaptic membrane may have tens to thousands of transmitter-gated ion channels, and the amount of transmitter released determines how many of these channels will be activated. The size of an EPSP will be a multiple of the number of vesicles, each containing several thousand transmitter molecules.
An inhibitory postsynaptic potential (IPSP) is a hyperpolarization that makes the membrane potential more negative and pushes the neuron away from its excitation threshold. At most inhibitory synapses, IPSPs are produced when neurotransmitters like GABA or glycine bind to receptors and cause negative chloride ions to enter the cell. When an inhibitory synapse is closer to the soma than an excitatory synapse, it can counteract positive current flow and decrease the size of the EPSP, a mechanism called shunting inhibition (Bear, Connors, & Paradiso, 2026). The balance between EPSPs and IPSPs at any moment determines whether a neuron fires, making this push-pull dynamic the fundamental language of the EEG.
Integrating Postsynaptic Potentials
Integration is the summation of EPSPs and IPSPs at the unmyelinated axon hillock, the neuron's decision-making junction.
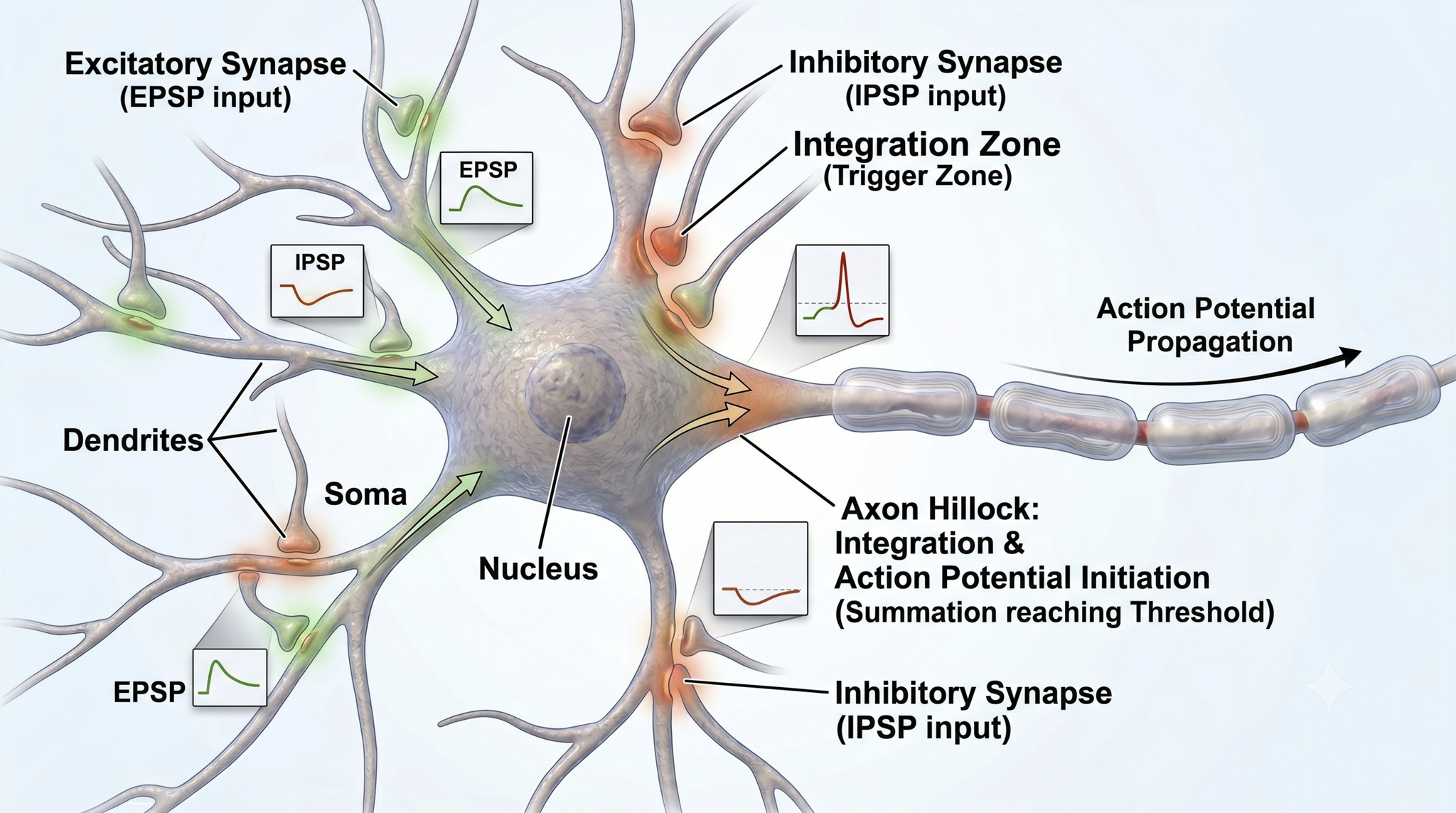
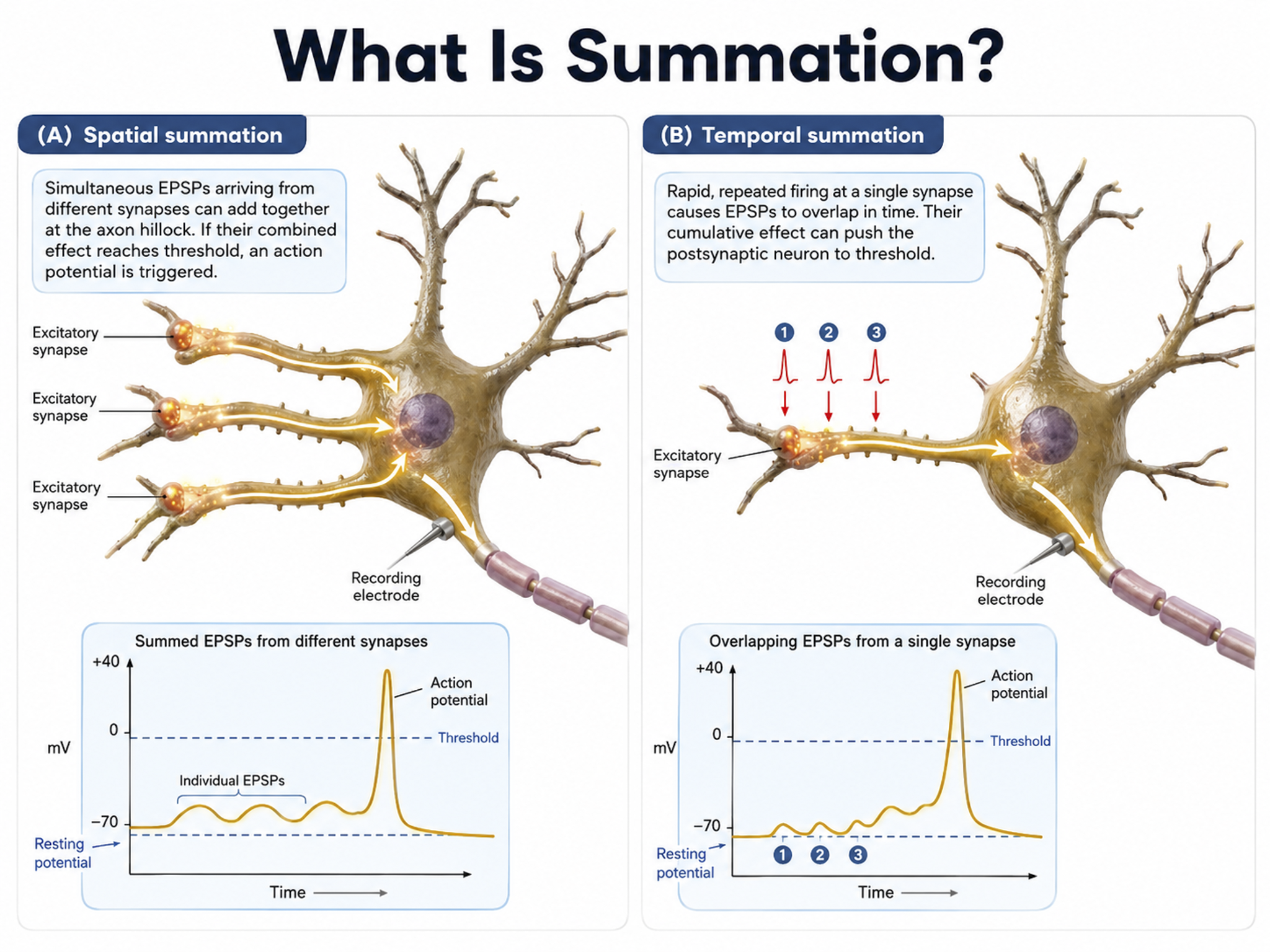
The axon hillock of a postsynaptic neuron uses two methods to sum EPSPs and IPSPs: spatial and temporal summation.
In spatial summation, the axon hillock sums the simultaneous postsynaptic potentials (PSPs) from thousands of synapses on dendrites.
In temporal summation, the axon hillock adds the PSPs from presynaptic neurons that repeatedly fire within a 1-15-ms time window.
Each EPSP depolarizes the axon hillock by about 0.5 mV. If there were no competing IPSPs, it would take about 30 EPSPs to trigger an action potential. Each IPSP hyperpolarizes the axon hillock by about 0.5 mV. If the summated EPSPs and IPSPs move the axon hillock from a resting potential of -70 mV to a threshold of excitation of -55 mV, sodium channels in the axon hillock membrane open, and an action potential propagates down the axon. Graphic adapted from © 2003 Josephine Wilson.
Check out the YouTube video, Best Action Potential Explanation.
Action Potentials
This section covers how action potentials transmit signals along axons and explains two key principles, the all-or-none law and the rate law, along with the two modes of conduction: unmyelinated and myelinated. Understanding conduction speed and efficiency explains why demyelinating diseases like multiple sclerosis are so devastating.
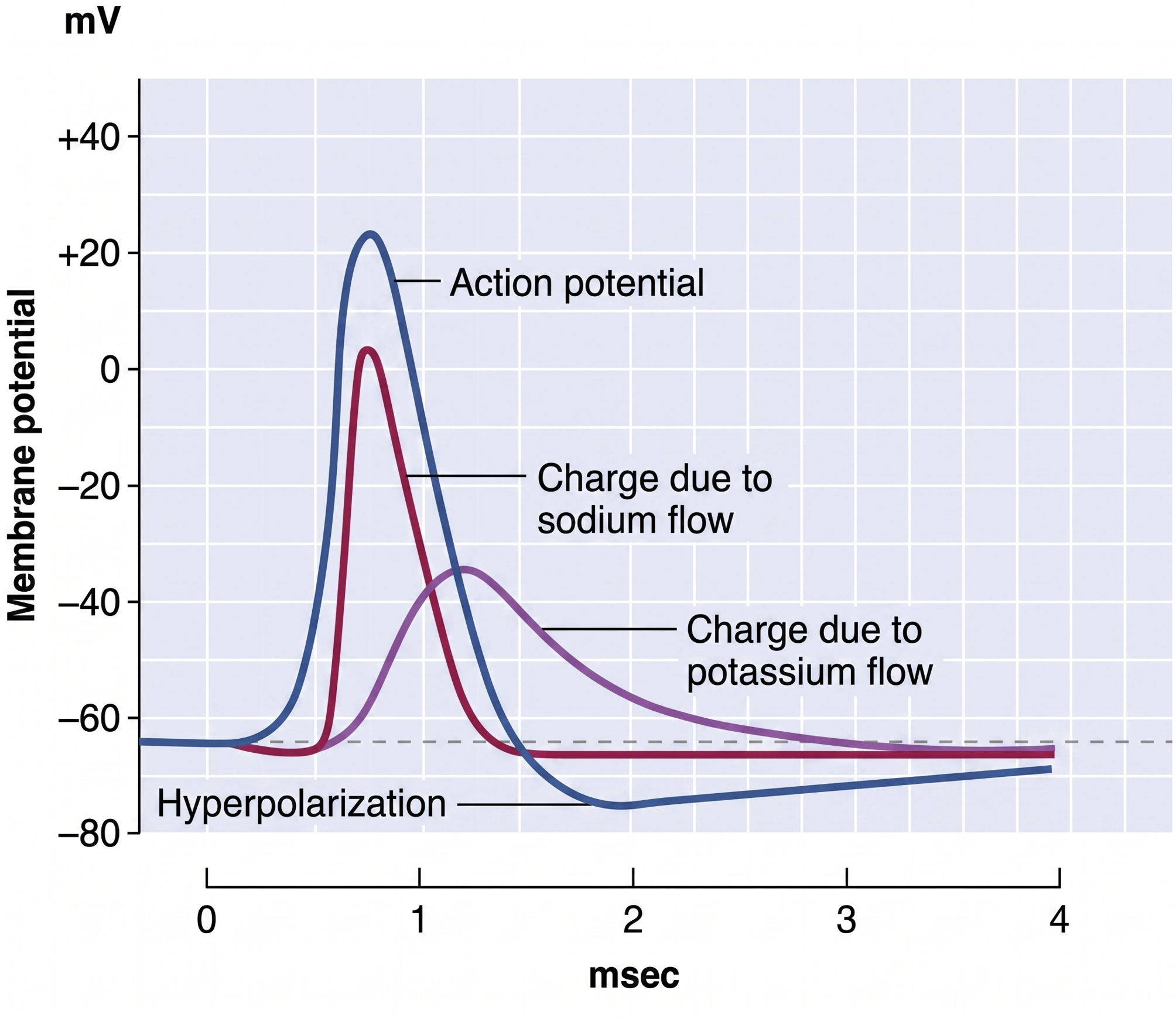
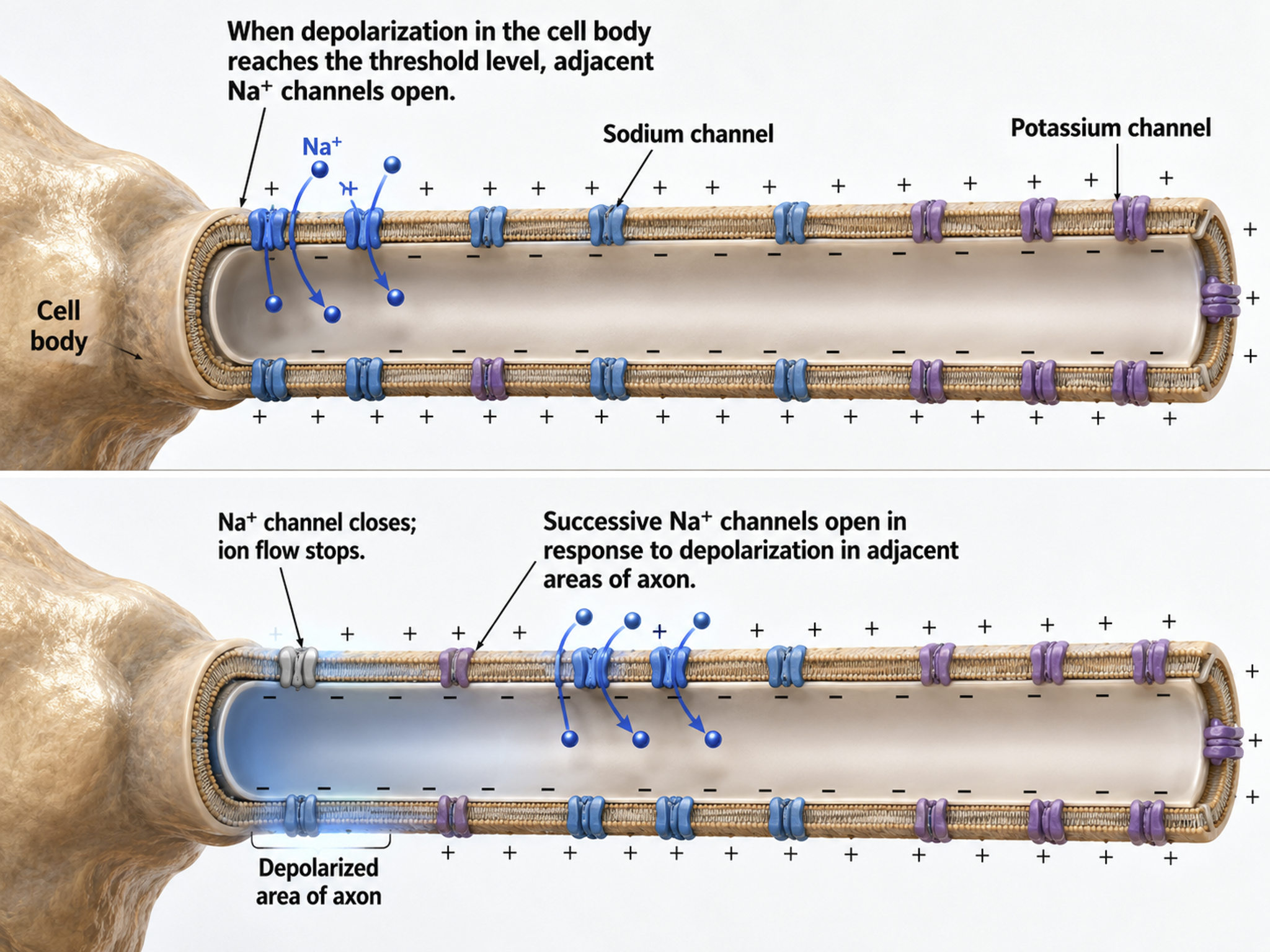
An action potential is a brief electrical impulse that transmits information from the axon hillock to the terminal button. This wave of positive charge only travels in one direction because the preceding segment is refractory due to the closing of its sodium channels. An action potential takes 1-2 ms from the point the axon hillock reaches its threshold to its repolarization to a negative resting potential.
The action potential unfolds as a rapid, self-reinforcing sequence of ion movements through voltage-gated ion channels, channels that open and close in response to changes in membrane voltage. When summation drives the axon hillock to threshold, voltage-gated sodium channels open and sodium rushes in, depolarizing the membrane toward +30 mV in the rising phase. Within about a millisecond these sodium channels inactivate while voltage-gated potassium channels open, allowing potassium to leave and repolarize the membrane in the falling phase. A brief overshoot of potassium efflux produces a short hyperpolarization before the resting potential is restored (Bear, Connors, & Paradiso, 2026).
Because the sodium channels remain inactivated during the absolute refractory period, the membrane cannot fire again no matter how strong the input, which forces the impulse to travel in one direction and sets an upper limit on firing rate. During the following relative refractory period, a stronger-than-usual input is required to fire the neuron, the mechanism that allows the rate law described below to encode stimulus intensity (Breedlove & Watson, 2023).
Action potentials travel down axons, which branch multiple times and terminate at synapses. The all-or-none law and rate law describe action potential transmission. The all-or-none law states that once an action potential is triggered in an axon, it is propagated, without decrement, to the end of the axon.
The rate law states that neurons represent the intensity of a stimulus by variation in the rate of axon firing. More intense stimuli shorten the interval before a neuron can fire again, allowing it to fire more rapidly; an intense stimulus can cause a neuron to fire every 2 or 3 ms, while a weak stimulus might lengthen the time lag to every 4 or 5 ms. Action potential graphic © extender_01/Shutterstock.com.
We can compare action potential conduction to the movement of water through a leaky garden hose.
Garden hose: water can take two paths, inside the hose or through holes in its wall, and the majority of the water will flow where movement is easiest. For a small-diameter hose with many large holes, most of the water will travel through the leaks. Conversely, for a large-diameter hose with only a few small holes, the bulk of the water will remain inside.
Axon: positive charge can take two paths, inside the axon or through pores in its membrane. Like water, a positive charge will take the path of least resistance. For a small-diameter axon with many open sodium ion channels, the majority of the current will exit the axonal membrane to the extracellular fluid. Small diameter, unmyelinated axons transmit action potentials without weakening since sodium ion channels constantly regenerate this signal. This method is slow because the signal travels step-by-step, small segment by small segment, and waits for sodium channels to admit enough positive ions to reach the excitation threshold.
This method also consumes considerable energy since sodium-potassium transporters, powered by ATP, are located across the axon membrane to exchange three sodium for two potassium ions.
Conversely, for a large-diameter axon with few open ion channels, the bulk of the current will remain inside the axon's interior. Wider spacing between adjacent ion channels means that the action potential can depolarize a longer axon segment, which increases conduction velocity (Bear, Connors, & Paradiso, 2026).
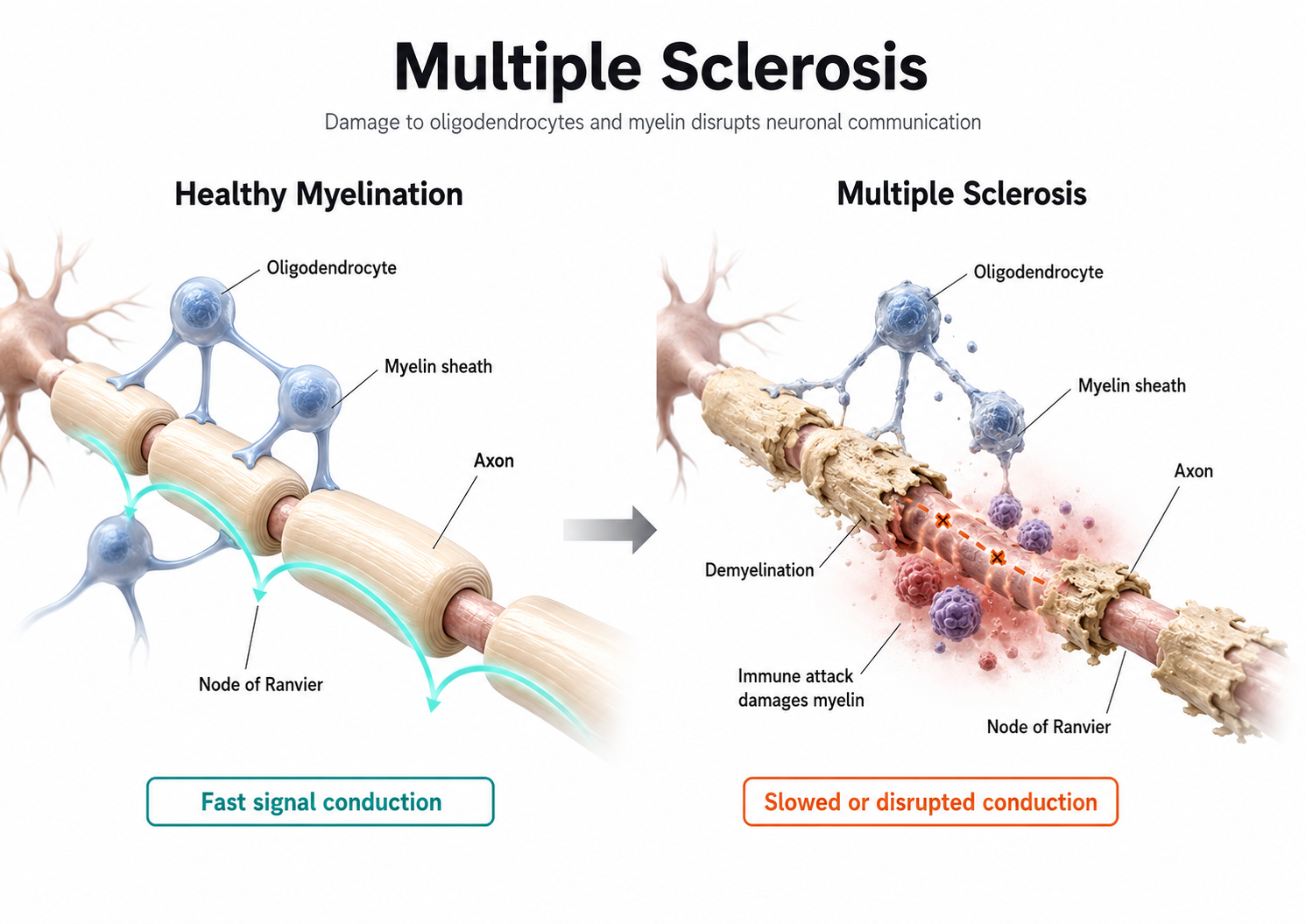
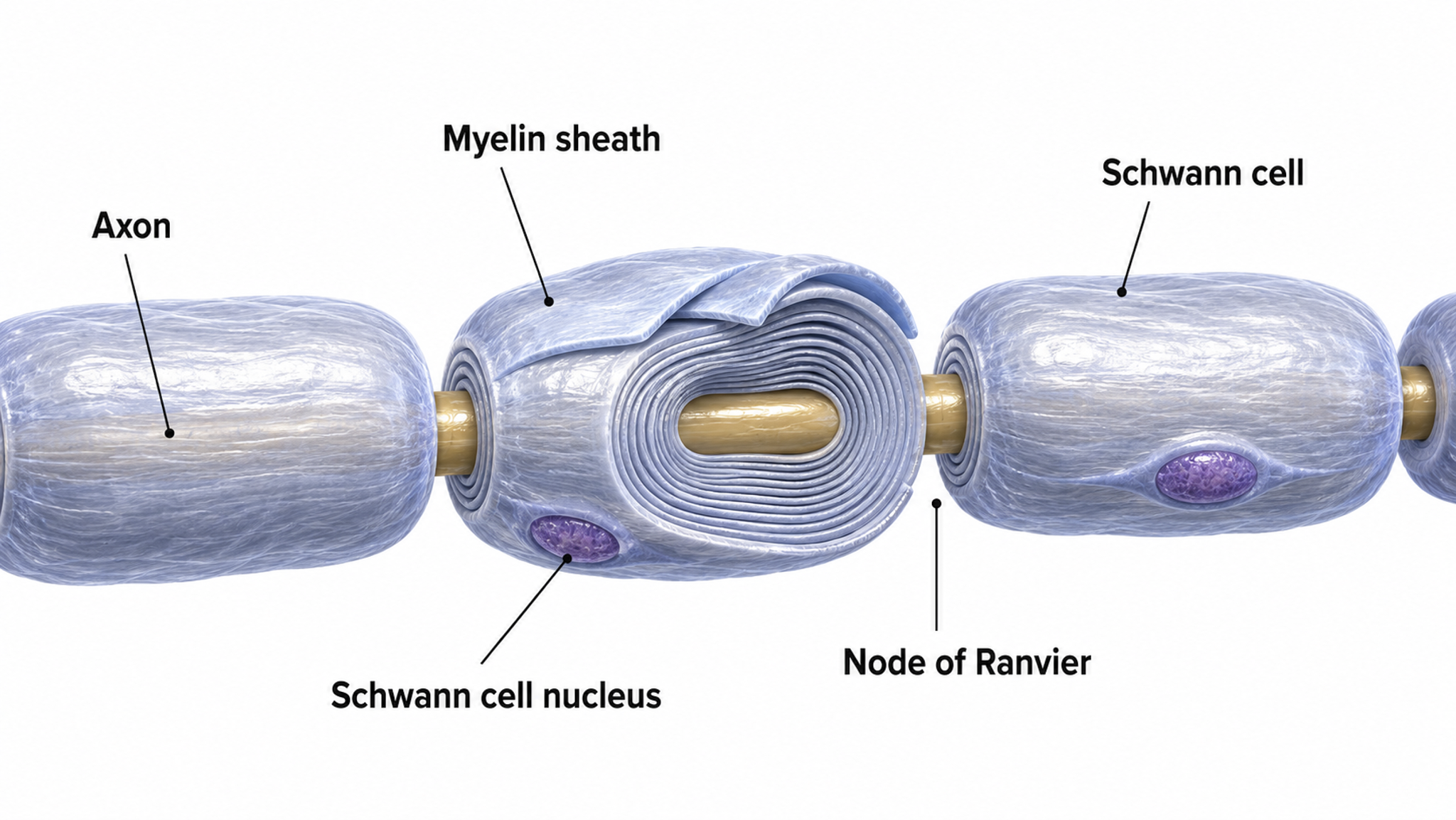
Medium-to-large diameter myelinated axons transmit action potentials using a method called saltatory conduction. Each segment of insulating myelin is almost 1-mm long, while the gaps between segments, called nodes of Ranvier, are 1 to 2 thousandths of a millimeter. An action potential weakens under each myelinated segment (cable properties) and is then regenerated at each Ranvier node. The destruction of this insulation by demyelinating diseases like multiple sclerosis (MS) can be devastating because it disrupts neuron-to-neuron communication.
Saltatory conduction can be 200 times faster because the action potential jumps from node to node in 1-mm steps, instead of steps that are a thousand times smaller. This method is also more energy-efficient because sodium-potassium transporters are only needed at the nodes of Ranvier, where ion exchange is possible. These transporters account for about 40% of a neuron's energy expenditure (Breedlove & Watson, 2023; Garrett, 2003).
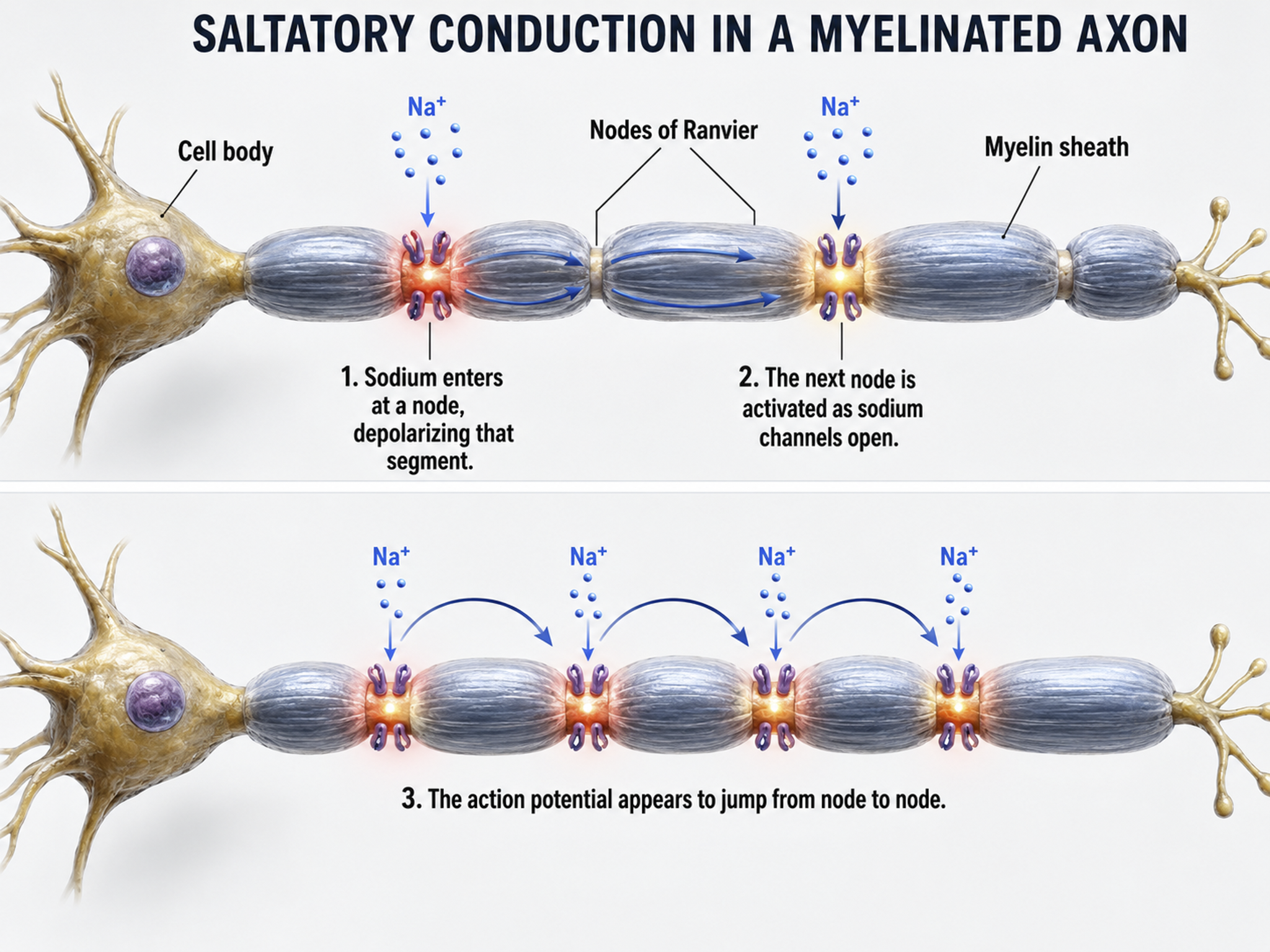
In summary, the type of neuron, the presence or absence of myelin, and the integrity of that myelin sheath all determine how quickly and efficiently neural signals travel, factors that directly affect the EEG patterns clinicians observe during assessment and training.
Key Takeaways
Neurons come in three functional classes, sensory, motor, and interneurons, and they work in partnership with glial cells that actively modulate signaling rather than merely supporting it. Each neuron maintains a resting membrane potential of about -70 mV that the sodium-potassium pump preserves at high metabolic cost. Graded excitatory and inhibitory postsynaptic potentials summate at the axon hillock, and when they reach the threshold of excitation an all-or-none action potential travels down the axon. Myelination and axon diameter determine how quickly and efficiently that signal propagates, which is why demyelinating disease disrupts communication. These postsynaptic potentials, not the action potentials, are the primary source of the scalp EEG.
Check Your Understanding
- How do sensory neurons, motor neurons, and interneurons differ in the direction and purpose of the information they carry?
- Why does the sodium-potassium pump make neurons such heavy energy consumers, and what happens to the EEG when a neuron's energy supply is compromised?
- How do spatial and temporal summation at the axon hillock determine whether a neuron reaches its threshold of excitation?
- Why does saltatory conduction in myelinated axons increase both speed and energy efficiency compared with conduction in unmyelinated axons?
- Why does the scalp EEG reflect postsynaptic potentials rather than action potentials?
Synaptic Transmission
This section explains how neurons communicate across chemical synapses, including neurotransmitter co-release, extra-synaptic transmission, modulation, and the major neurotransmitter families and pathways. These mechanisms are central to understanding how the brain generates the electrical activity that neurofeedback clinicians measure and train.
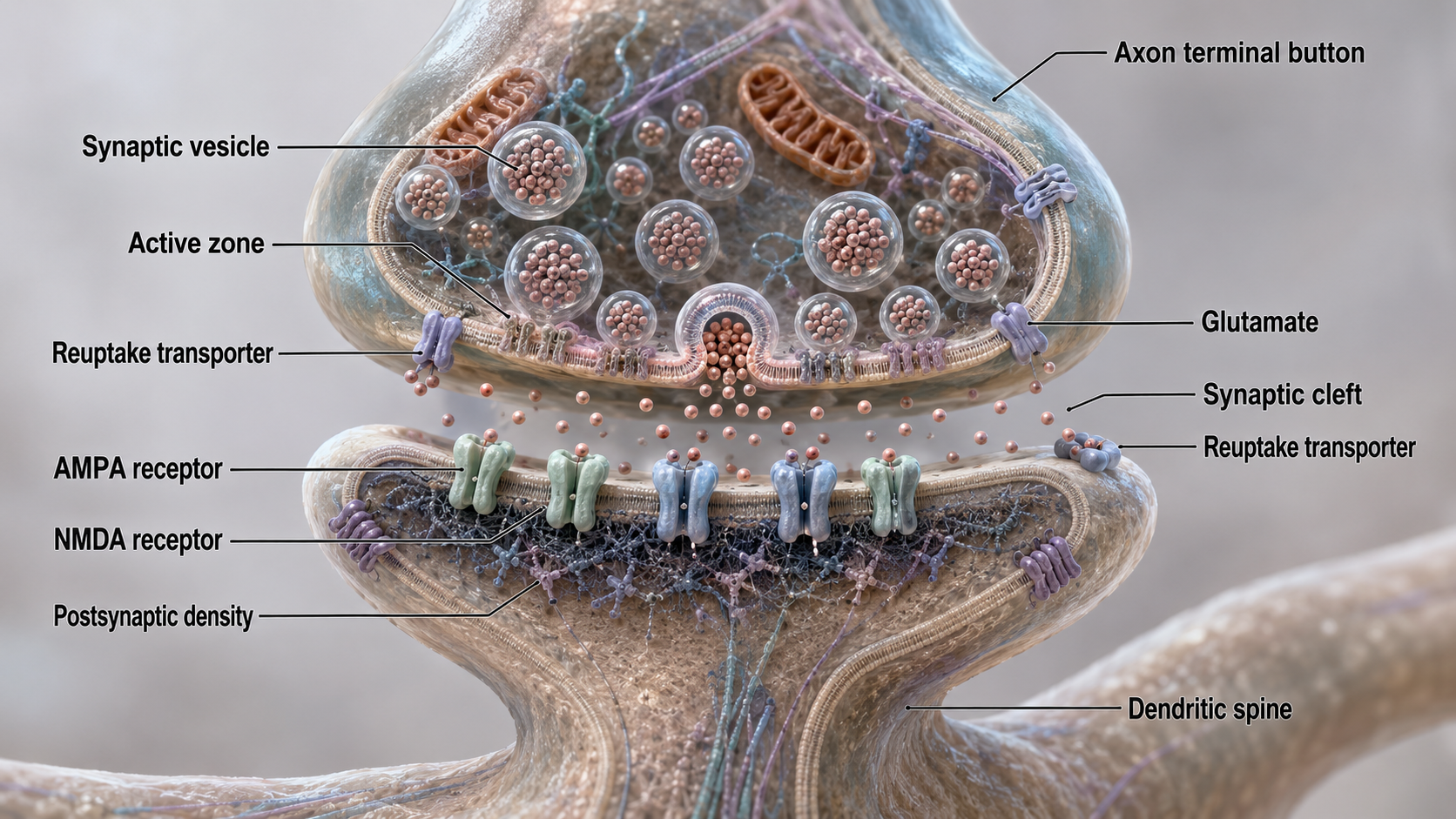
Neurons communicate through the release of over 200 neurochemicals and ions. Axon terminal buttons release neurochemicals across a 20-50-nm fluid-filled gap between presynaptic and postsynaptic structures called a synaptic cleft and into the extracellular fluid surrounding the neuron (Bear et al., 2020). Chemical synapses produce short-duration (millisecond) and long-duration (seconds to days) changes in the nervous system. Synapse animation without sound © 3Dme Creative Studio/Shutterstock.com.
Chemical synapses are functionally asymmetrical because the presynaptic neuron sends a chemical message and the postsynaptic neuron receives it. They are structurally asymmetrical because the presynaptic element (axon) contains vesicles containing NTs, and the postsynaptic element (dendrite) does not. NT release from a terminal button is called exocytosis (Breedlove & Watson, 2023). Synapse graphic adapted from © SciePro/Shutterstock.com.
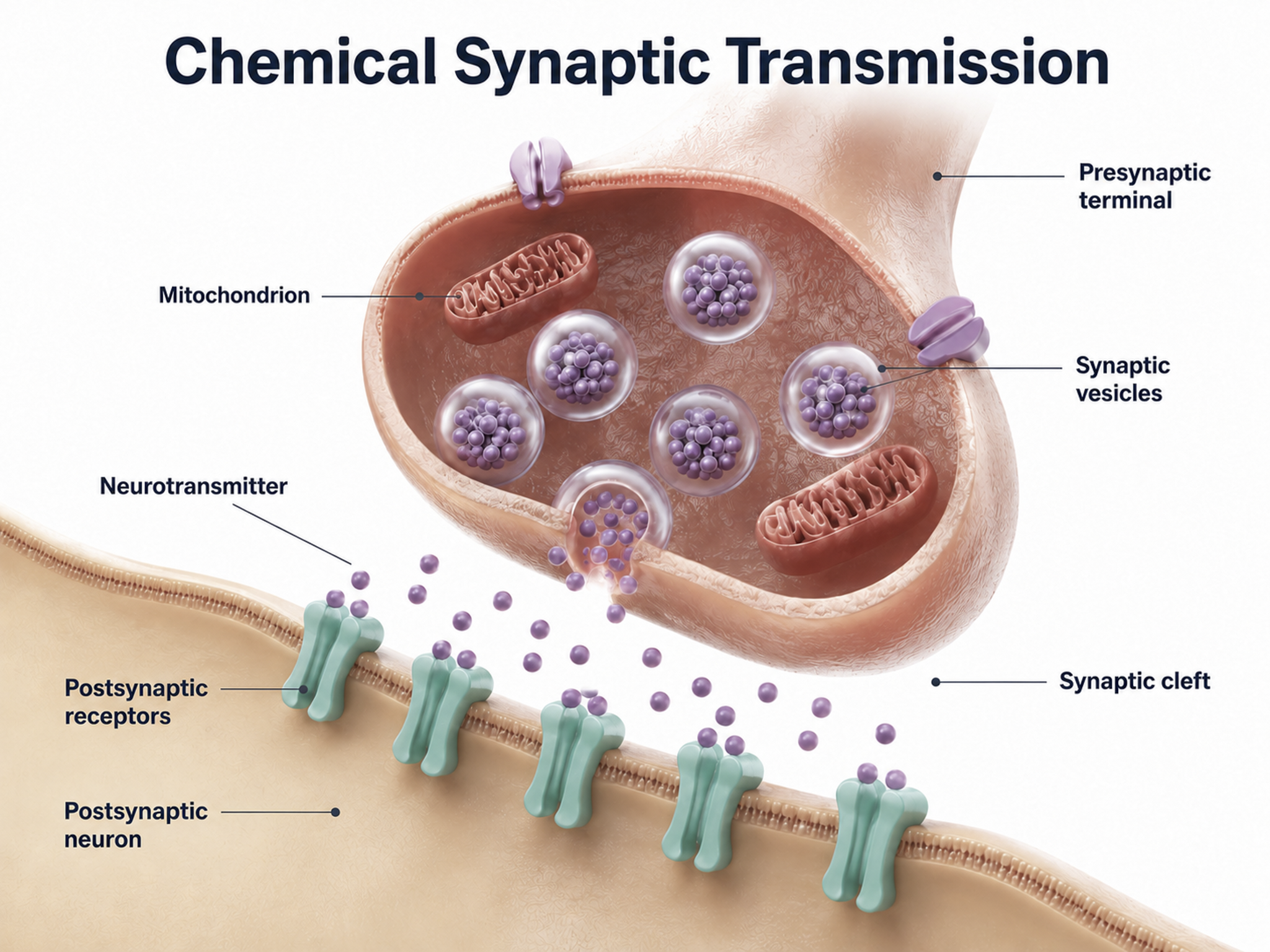
In the graphic below, an axon terminal button releases NTs into the synaptic cleft. NTs briefly bind to receptors on a dendritic spine through reversible, noncovalent interactions and then disengage after they initiate small graded potential changes (e.g., EPSPs or IPSPs) or more diverse, gradual, and long-lived actions (e.g., creating second messengers inside the target neuron). For clinicians, this distinction matters: fast ionotropic effects contribute to the moment-to-moment EEG, while slower metabotropic effects drive the long-term changes associated with neurofeedback training outcomes.
Ionotropic receptors open an ion pore directly. When a neurotransmitter binds to the receptor, the channel opens and ions flow rapidly across the membrane, producing fast and short-lived effects. Metabotropic receptors act more slowly. Instead of opening a channel themselves, they trigger a sequence of metabolic steps that activate a second messenger system inside the neuron, producing slower and longer-lasting changes. Chemical synapse graphic © nobeastsofierce/Shutterstock.com.
.jpg)
Neurotransmitter Co-Release
Old-school view: according to Dale's law, a neuron can only release one NT at a synapse.
New-school view: neurons can release a classical NT and a peptide.
Dale's law proposed that a neuron releases only one NT. However, researchers have found increasing evidence of NT co-release (Svensson et al., 2019). A neuron can store different NTs in separate types of vesicles (Hökfelt et al., 2003). Neurons can also store multiple NTs in the same vesicles (e.g., ATP and glutamate), although they may not release them simultaneously (Merighi et al., 2011; Xia et al., 2009). Co-release adds a layer of complexity to synaptic communication; a single neuron can send multiple chemical messages, producing nuanced effects on its postsynaptic partners.
![]()
Extra-Synaptic Transmission: Think Outside the Cleft
This section covers three mechanisms by which neurons release neurotransmitters outside of classical synapses: volume transmission, axonal varicosities, and retrograde transmission. These extra-synaptic pathways help explain how neurotransmitters influence broad regions of the brain, not just the neurons immediately across a synaptic cleft.
Neurons release NTs outside of classical synapses. These mechanisms include release from terminal buttons into the extracellular space, axonal varicosities, and retrograde transmission.
Old-school view: axon terminals only release NTs into the synaptic cleft.
New-school view: NT release also occurs outside of the synaptic cleft. Axonal varicosities (swellings in axon walls), dendrites, and the terminal button can release NTs into the extracellular space. Graphic © 3Dme Creative Studio/Shutterstock.com.
Volume Transmission
Volume transmission involves NT release and eventual binding to a receptor outside the synaptic cleft (Coggan et al., 2005). Do not confuse this process with volume conduction, which is the spread of an electrical signal when measured at some distance from its source. Graphic adapted from the American Scientist.
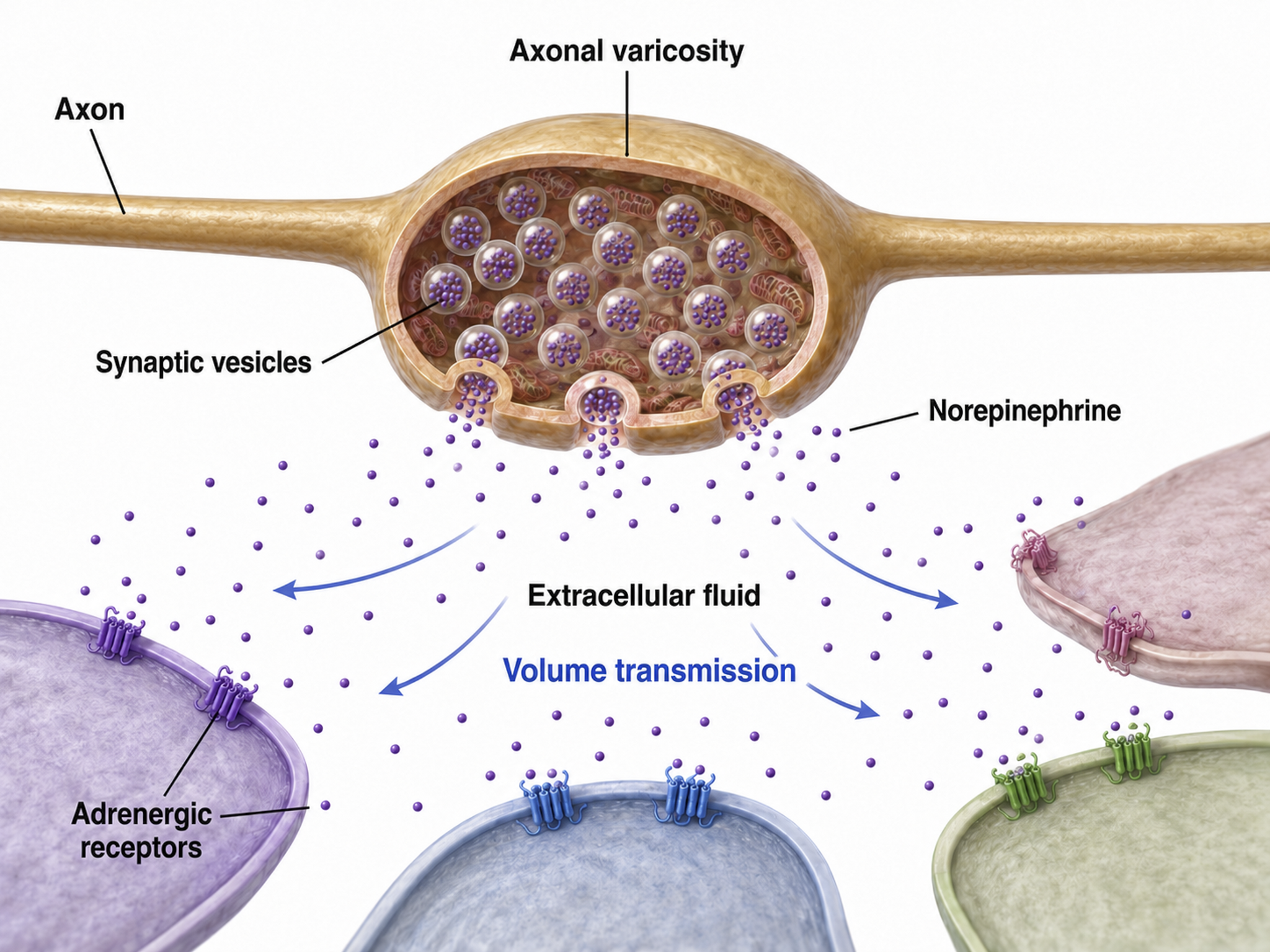
Axonal Varicosities
Axons can release NTs into the extracellular space through varicosities (swellings) along their length, analogous to drip irrigation (Breedlove & Watson, 2023).
Most neurons that release norepinephrine do not do so through terminal buttons on the ends of axonal branches. Instead, they usually release them through axonal varicosities, beadlike swellings of the axonal branches (Carlson & Birkett, 2019, pp. 82-83).
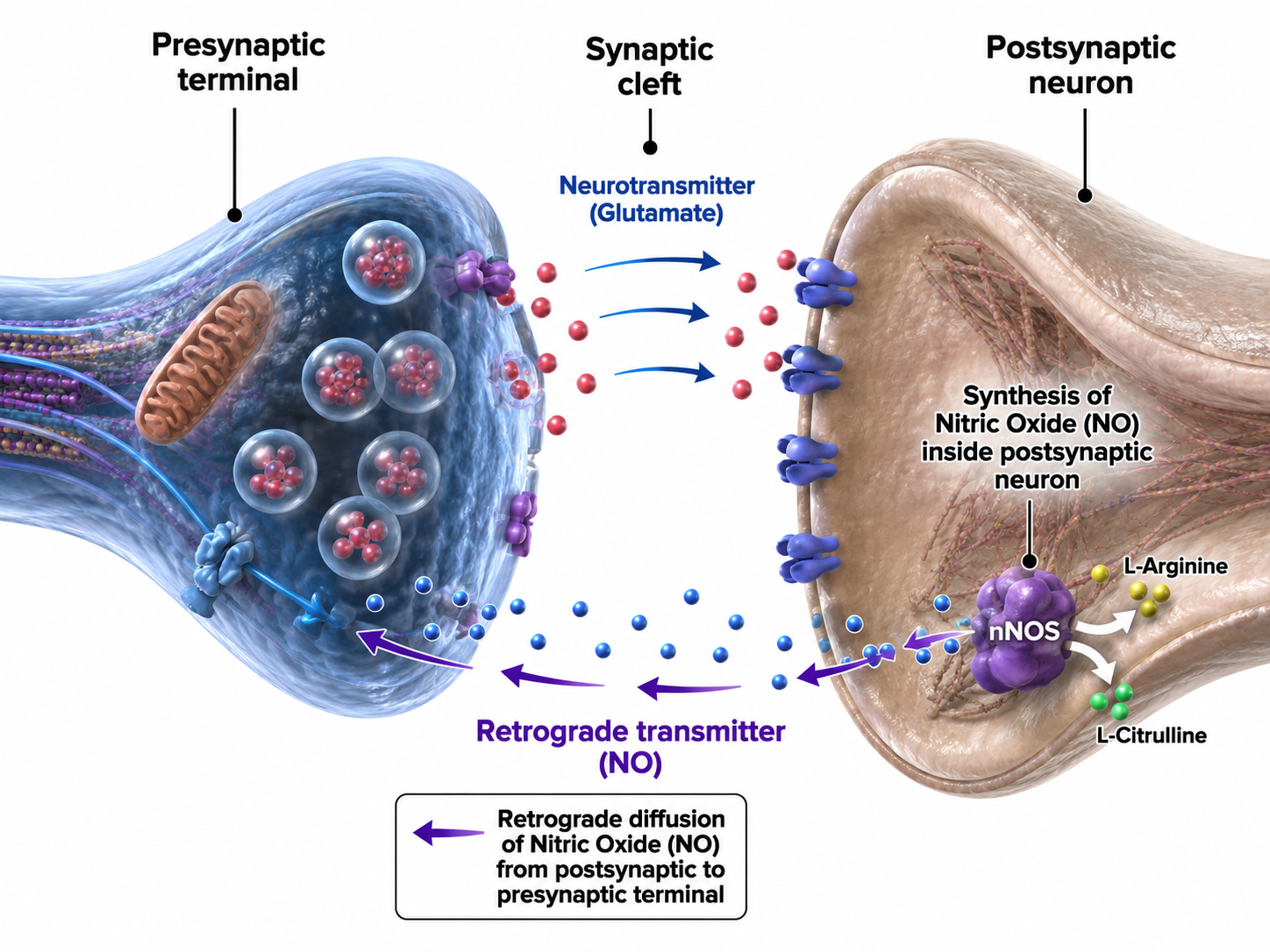
Retrograde Transmission
In retrograde transmission, a presynaptic neuron sends a chemical message to the postsynaptic neuron. In response, the postsynaptic neuron synthesizes and distributes an endocannabinoid (e.g., anandamide) or gas (e.g., nitric oxide, or NO) to the presynaptic neuron and its immediate active neighbors. Neurons synthesize these NTs on demand since they cannot be contained by vesicles. This backward signaling allows the postsynaptic neuron to fine-tune the messages it receives, a feedback loop that plays an important role in synaptic plasticity.
. . . this gaseous signal has a range of influence that extends well beyond the cell of origin, diffusing a few tens of micrometers from its site of production before it is degraded. This property makes NO a potentially useful agent for coordinating the activities of multiple cells in a localized region and may mediate certain forms of synaptic plasticity that spread within small networks of neurons (Purves, 2017, pp. 142-143).
Retrograde NTs can bind to membrane-bound receptors or diffuse into the target cell, initiating second messenger production to adjust synaptic efficiency in learning and memory (Breedlove & Watson, 2023).
Modulation
This section explains how the nervous system fine-tunes its signals through modulation of neurotransmitter release and receptor activity. Modulation is analogous to a volume control knob on a stereo preamplifier rather than a simple on/off switch; it provides analog adjustment rather than digital switching.

We will consider two of countless modulation mechanisms: modulation of NT release and modulation of NT action at its receptor.
Neurotransmitter Release Modulation
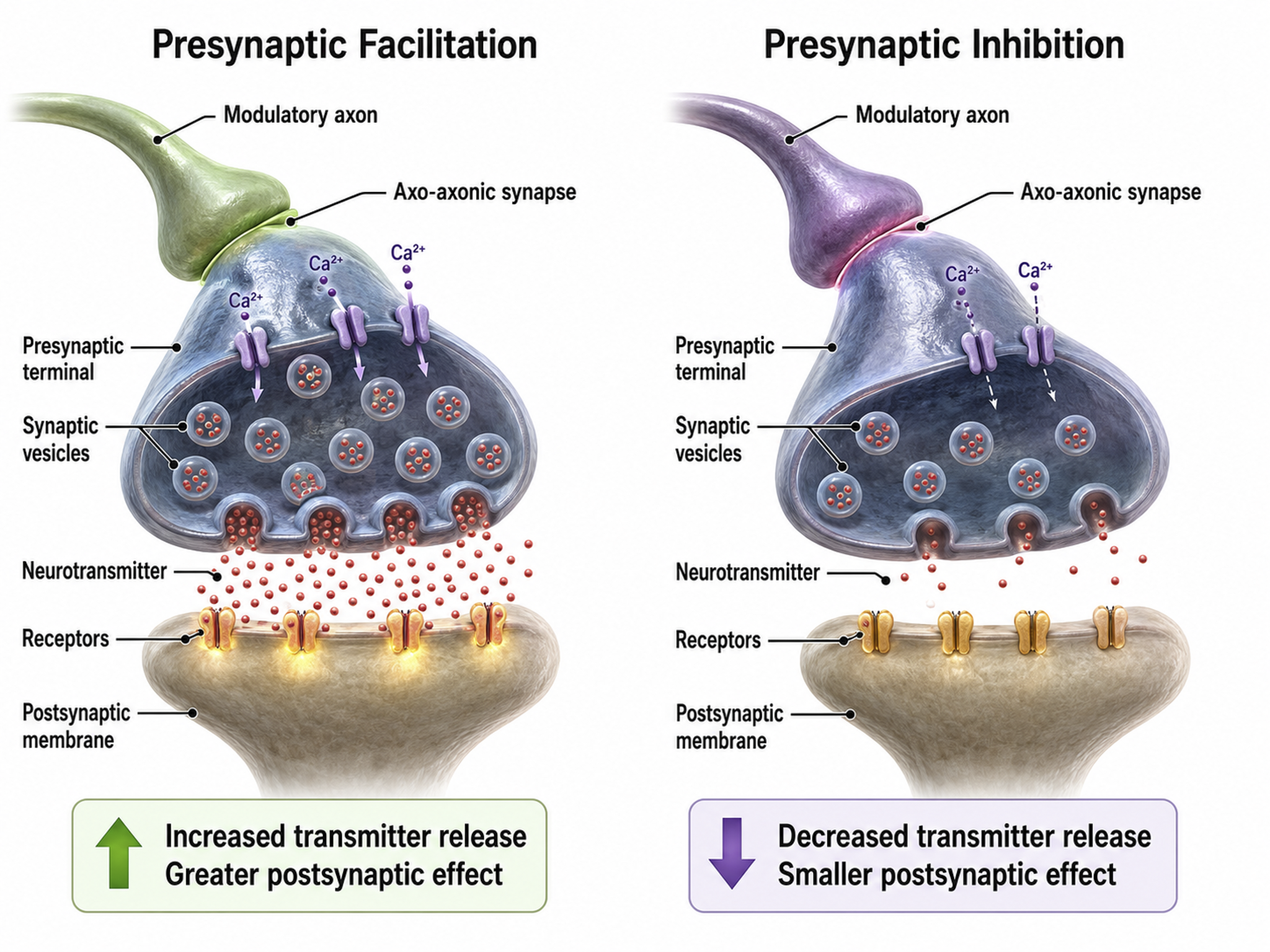
Axons can influence the amount of NTs released when an action potential arrives at an axon terminal through axoaxonic synapses (junctions between two axons).
Axoaxonic synapses do not affect the generation of an action potential, only the amount of neurotransmitter distributed. In presynaptic facilitation, a neuron increases the presynaptic neuron's neurotransmitter release by delivering a neurotransmitter that increases calcium ion entry into its terminal button. In presynaptic inhibition, a neuron decreases neurotransmitter release by reducing calcium ion entry. These modulatory effects are confined to a single synapse (Breedlove & Watson, 2023).
Autoreceptors Modulate Neurotransmitter Release
Autoreceptors are metabotropic receptors on the presynaptic membrane that function as a built-in feedback system. When NTs released into the synaptic cleft bind to autoreceptors, this hyperpolarizes the axon terminal button so it will release less NT when the next action potential arrives. This self-regulation prevents excessive neurotransmitter release, a mechanism that many psychotropic medications exploit.
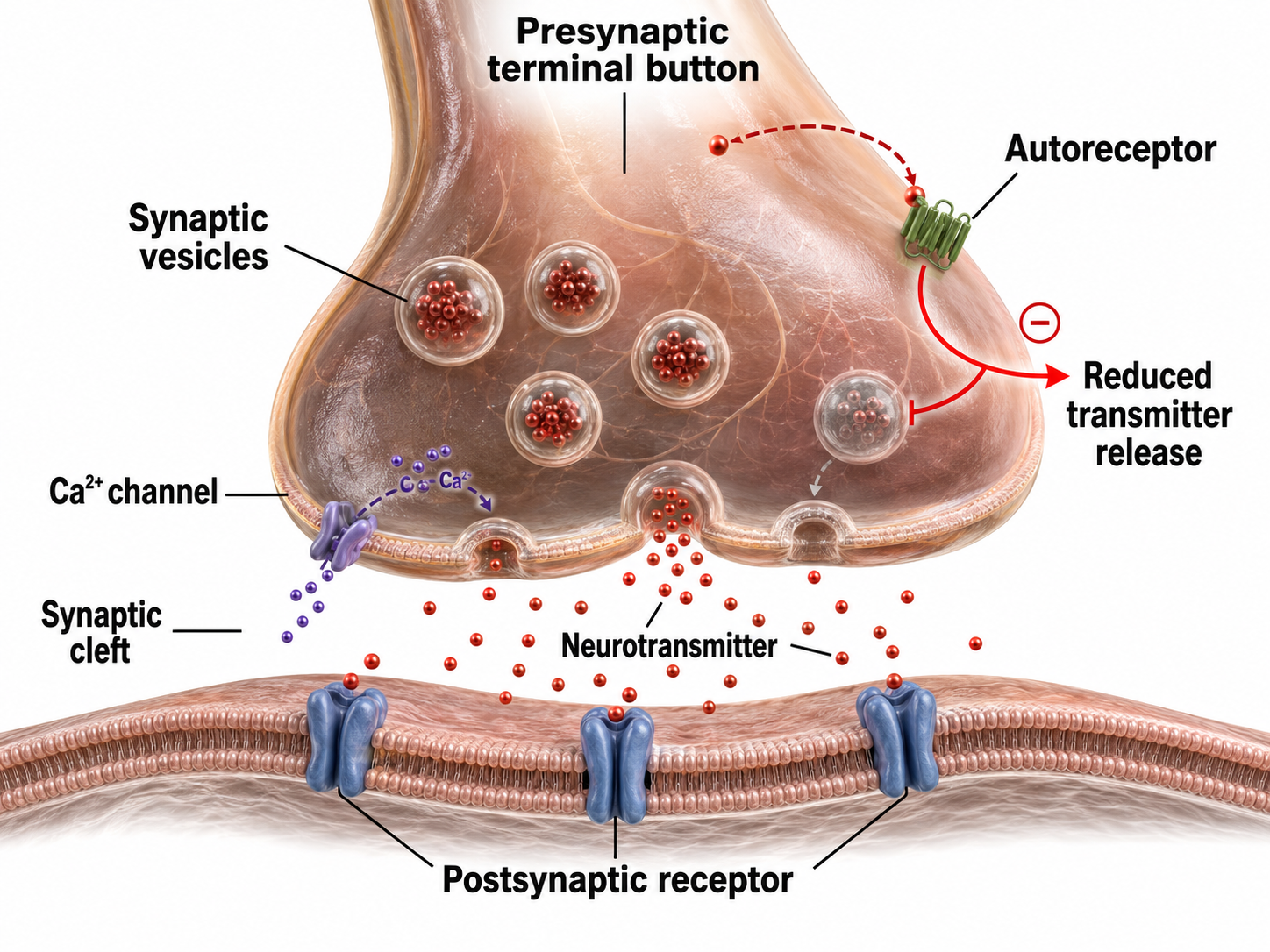
Neuromodulators Adjust Neurotransmitter Action
A postsynaptic neurotransmitter receptor contains two functionally distinct binding sites. The orthosteric site is the primary location where the neurotransmitter binds, and it behaves like an on-off switch that activates the receptor. The allosteric site is a separate, secondary location where modulators bind. Rather than switching the receptor on or off, it acts like a volume control, adjusting how strongly the receptor responds when a neurotransmitter occupies the orthosteric site.
Receptors contain binding sites for drugs like alcohol, ions, or endogenous molecules. When alcohol binds to its allosteric site, it strengthens GABA's binding at its orthosteric site, causing greater chloride entry into the neuron and increasing its hyperpolarization. Ingesting multiple CNS depressants (e.g., alcohol and barbiturates) can yield dangerous additive effects, amplifying GABA's action to a level that can depress or stop breathing. This is why clinicians conducting neurofeedback assessments should always inquire about substance use; CNS depressants will alter the EEG patterns observed during recording.
Types of Neurotransmitters
While the actual number of NTs is not known, more than 200 molecules have been identified. Each neurotransmitter may have multiple receptors, and a NT's effect, excitatory or inhibitory, depends on its interaction with specific receptors. The same NT can produce opposite results at different receptor subtypes (Breedlove & Watson, 2023).
The principal NT families include amino acid neurotransmitters (GABA, glutamate), amine neurotransmitters (acetylcholine, dopamine, serotonin), peptide neurotransmitters, also called neuropeptides (oxytocin, vasopressin), gas neurotransmitters or gasotransmitters (nitric oxide, carbon monoxide, hydrogen sulfide), purine neurotransmitters (ATP, adenosine), and lipid neurotransmitters (anandamide and 2-arachidonoylglycerol, or 2-AG). We adapted the table below from Breedlove and Watson (2023).
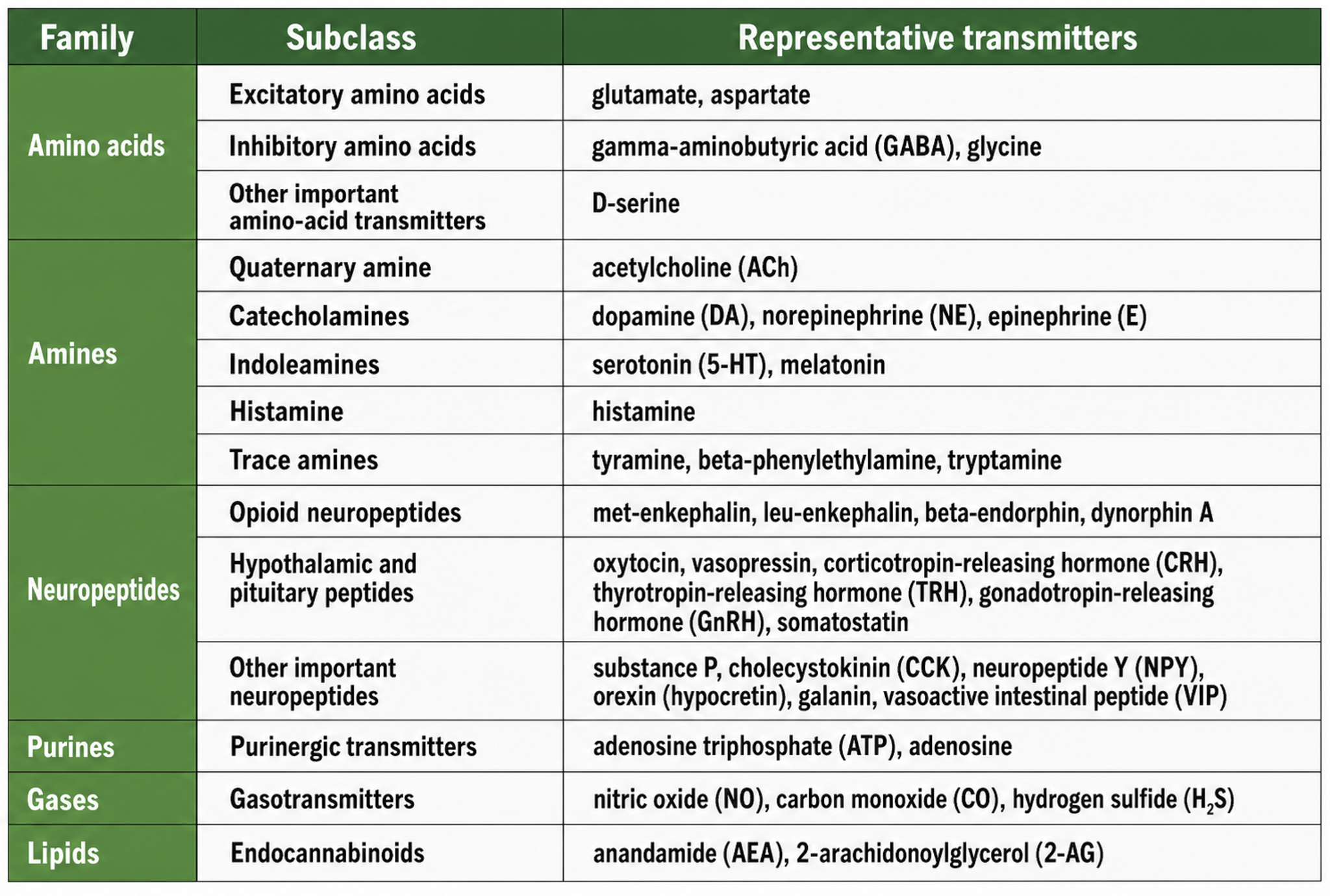
Neurotransmitter Pathways
Researchers have identified distinct pathways for acetylcholine, dopamine, norepinephrine, and serotonin. Understanding these pathways is critical for neurofeedback practitioners because they explain how specific neurotransmitter systems influence the EEG patterns targeted in training protocols. The reproduced diagrams are adapted from © Vasilisa Tsoy/Shutterstock.com.
Cholinergic pathways
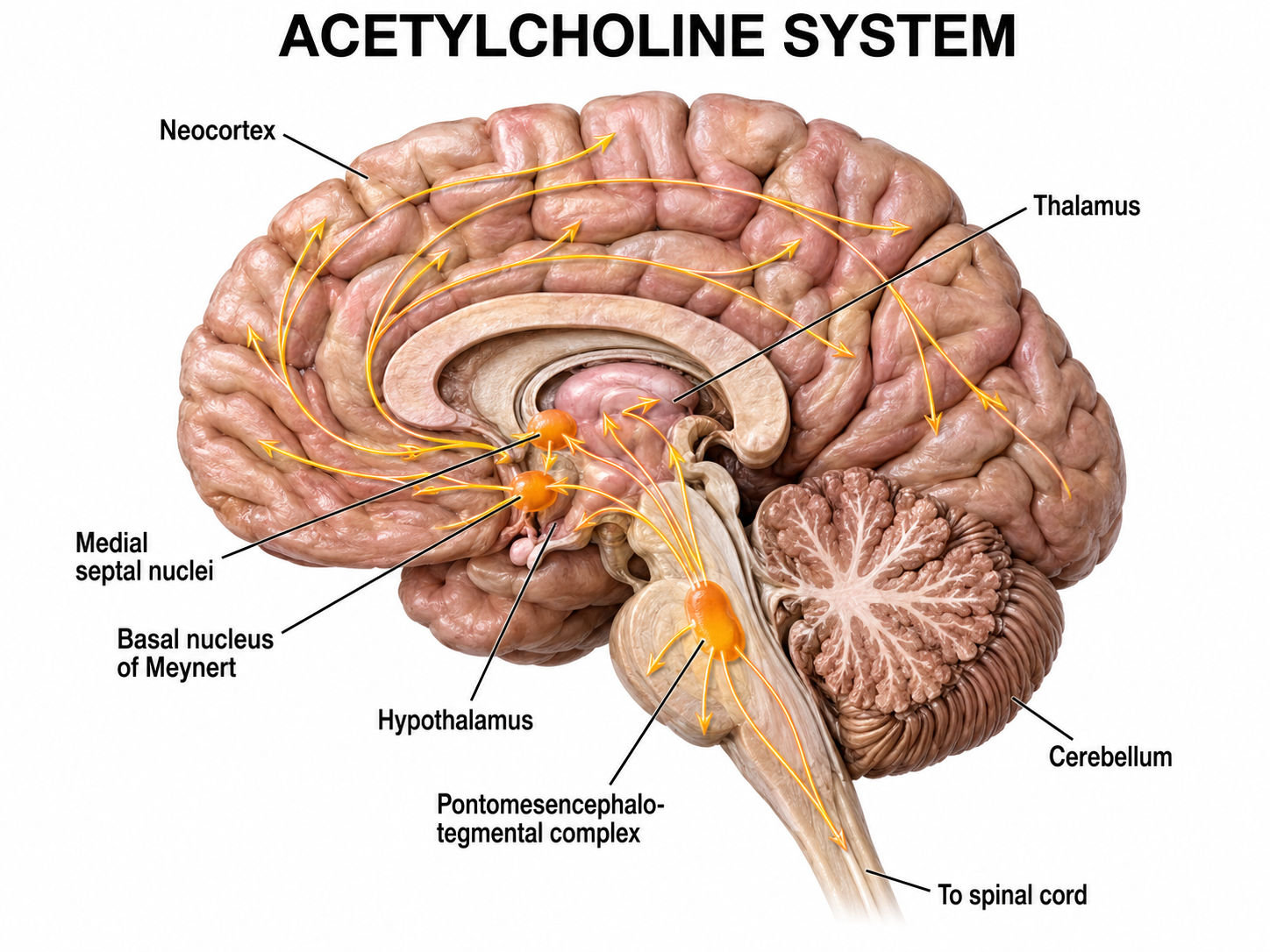
Cholinergic cell bodies and their projections originate in the basal forebrain and brainstem. Cholinergic pathways are involved in arousal, attention, memory, motivation, muscle contraction, and sleep.
Dopaminergic pathways
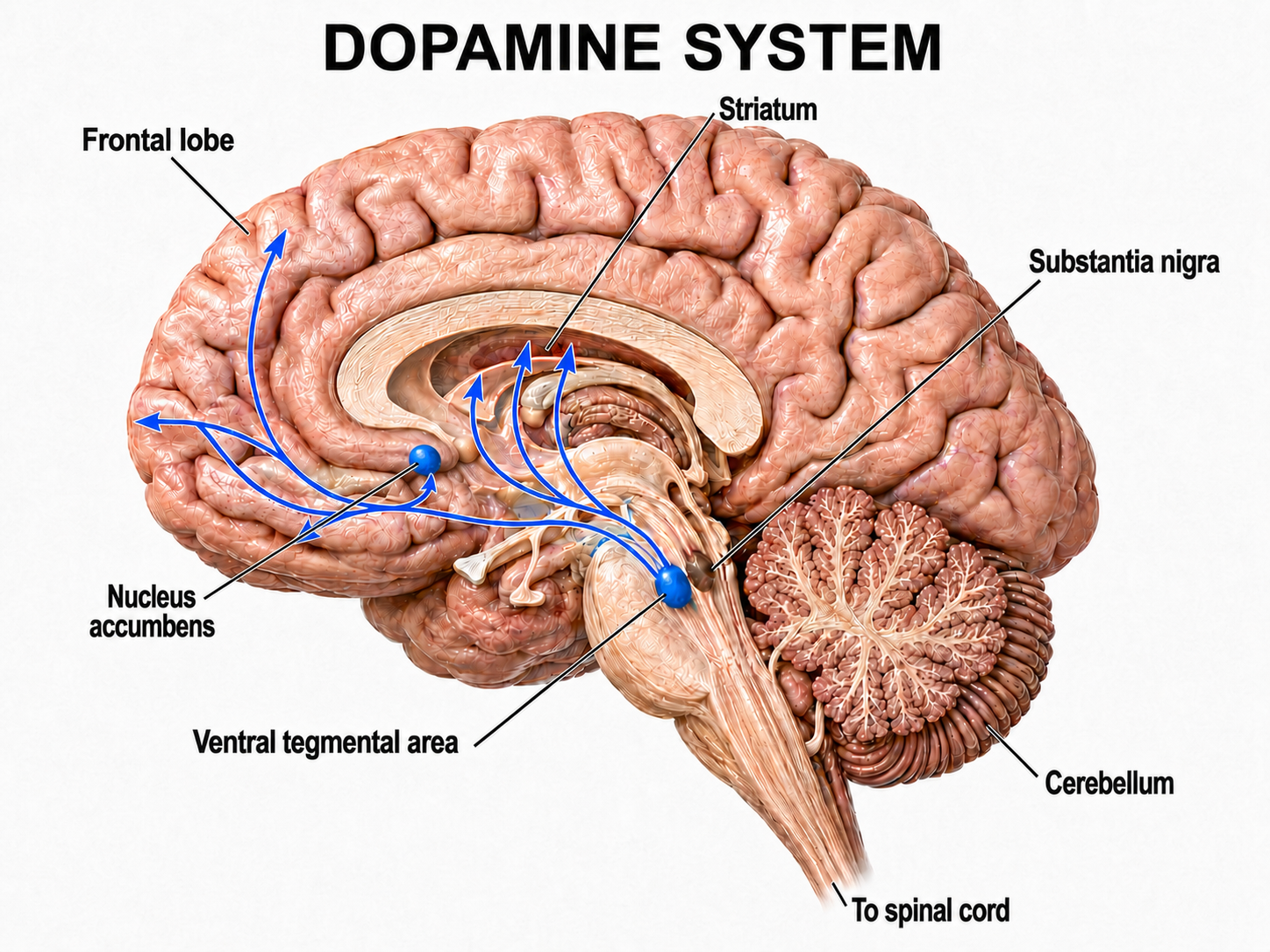
Two major dopaminergic pathways originate in the midbrain: the mesostriatal and mesolimbocortical pathways. Dopaminergic pathways are involved in addiction, motor control, and salience (reward- and threat-based motivation). These pathways are particularly relevant in neurofeedback because the dopamine reward system is engaged when clients receive positive feedback during training.
Noradrenergic pathways
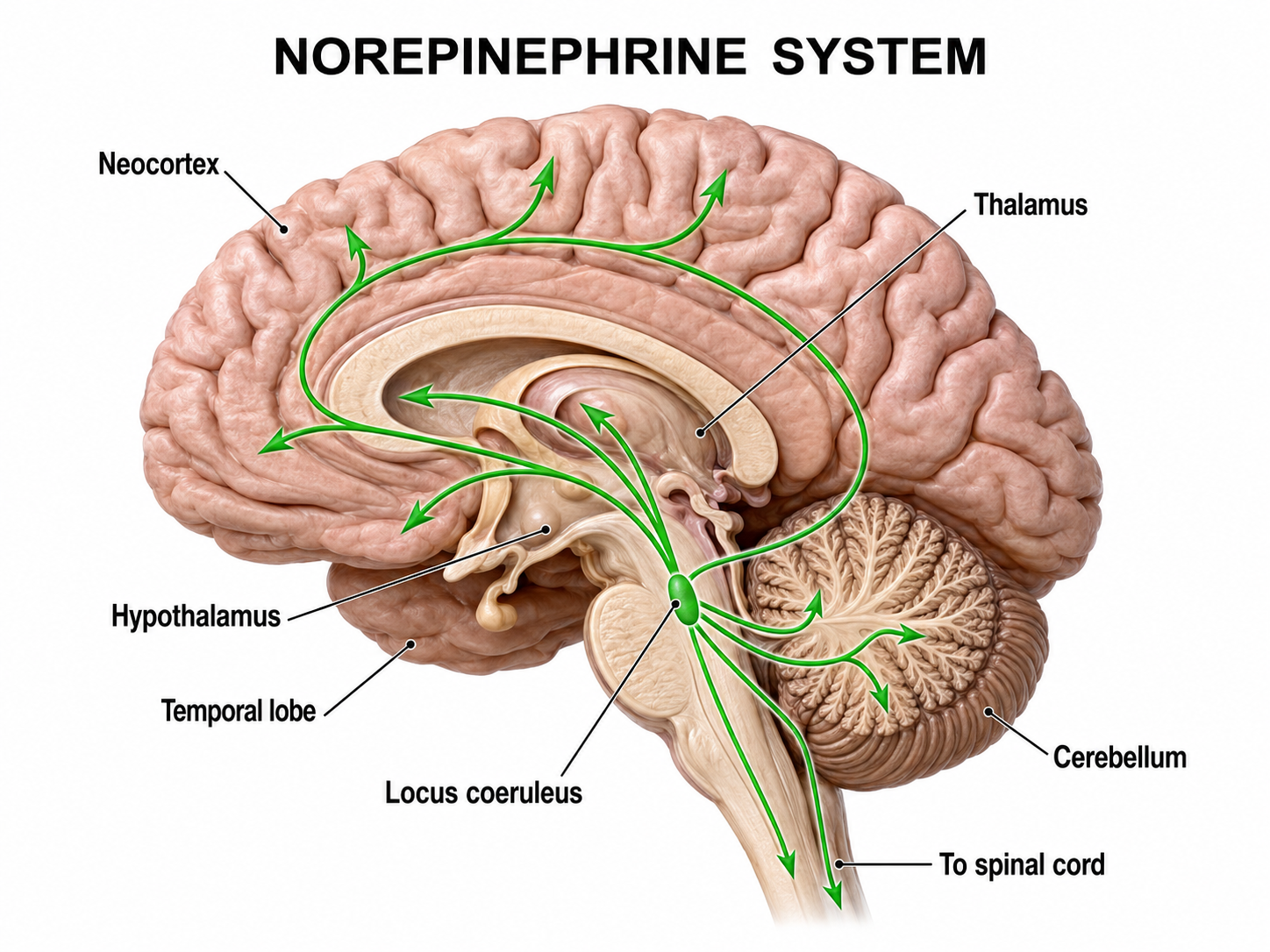
The noradrenergic pathways originate in the locus coeruleus of the dorsal pons and in the lateral tegmental cell groups of the pons and medulla. Noradrenergic pathways are involved in arousal, attention, memory, vigilance, sleep, and mobilizing the brain and body for action, including the fight-or-flight response. Dysregulation of these pathways is commonly seen in PTSD and anxiety disorders treated with neurofeedback.
Serotonergic pathways
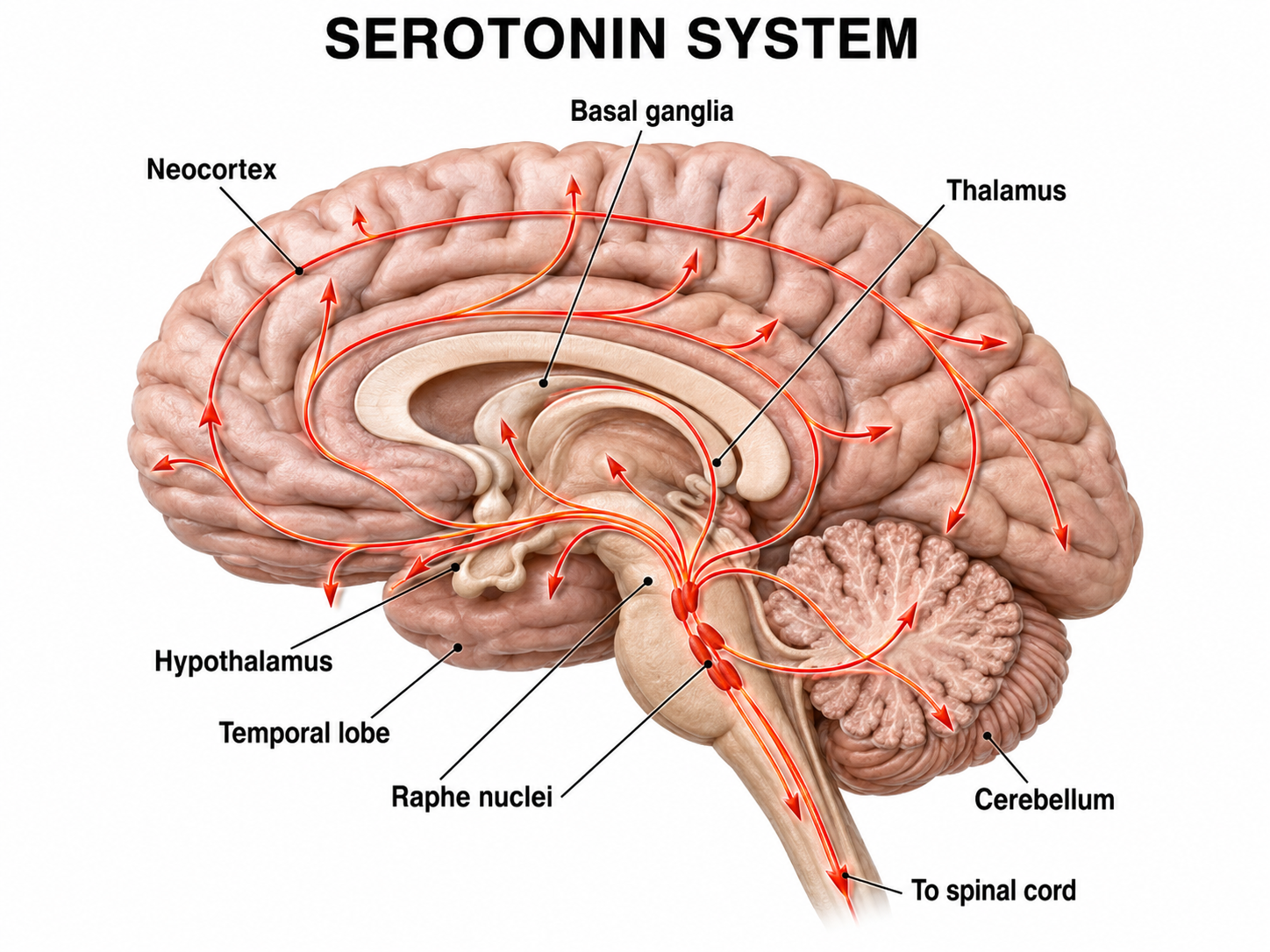
The serotonergic pathways originate in the brainstem and midbrain raphe nuclei. Serotonergic pathways are involved in appetite, mood, and sleep.
Termination of Neurotransmitter Action
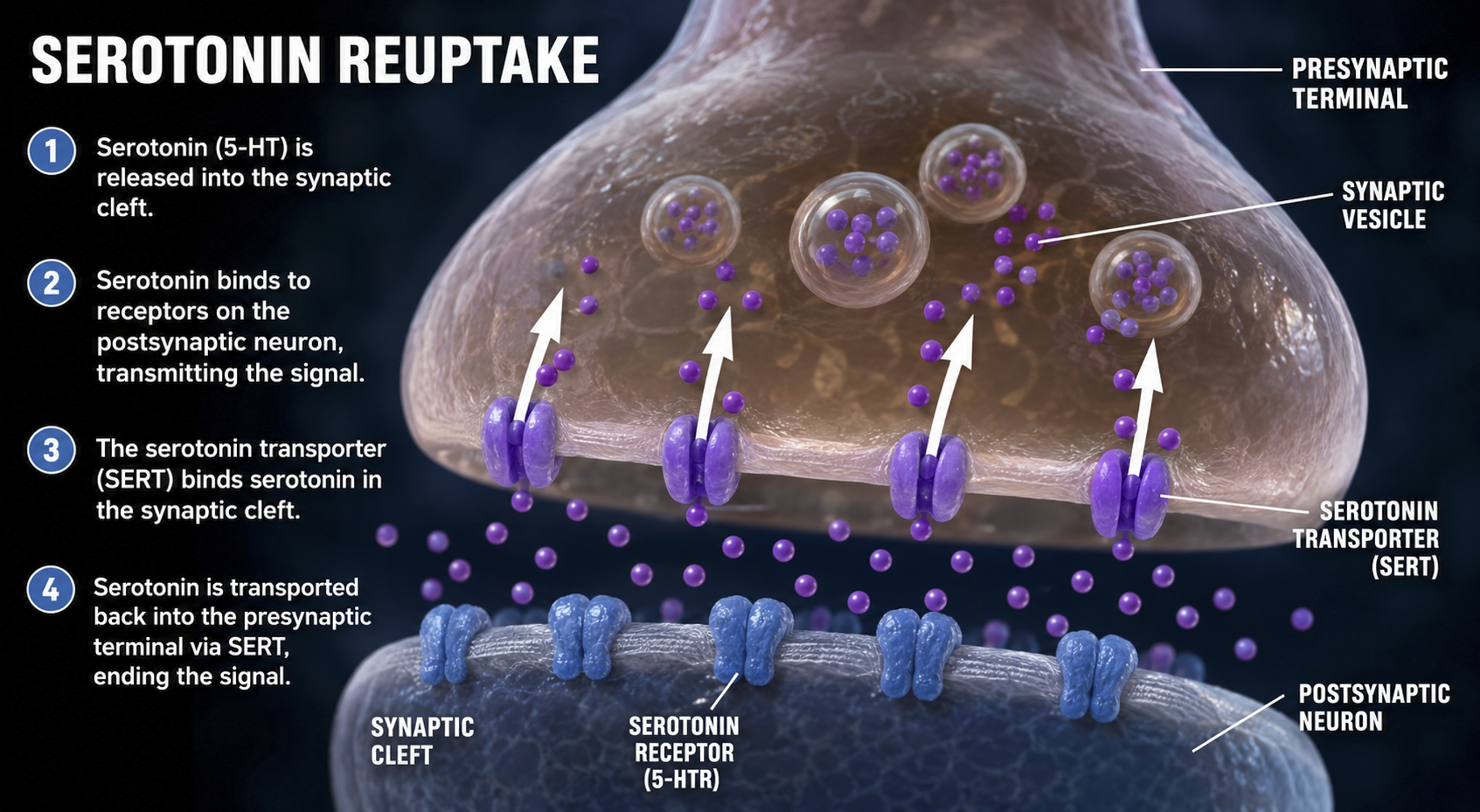
Following exocytosis, which is NT release from a presynaptic terminal vesicle, NT action is terminated by two main mechanisms: reuptake and enzymatic degradation. In reuptake, reuptake transporters located in the presynaptic terminal and astrocytes that enclose the synapse return NT molecules to the presynaptic neuron. This is the mechanism that SSRI antidepressants block; by inhibiting serotonin reuptake, they prolong serotonin's action in the synaptic cleft. Astrocytes remove glutamate from the synapse.
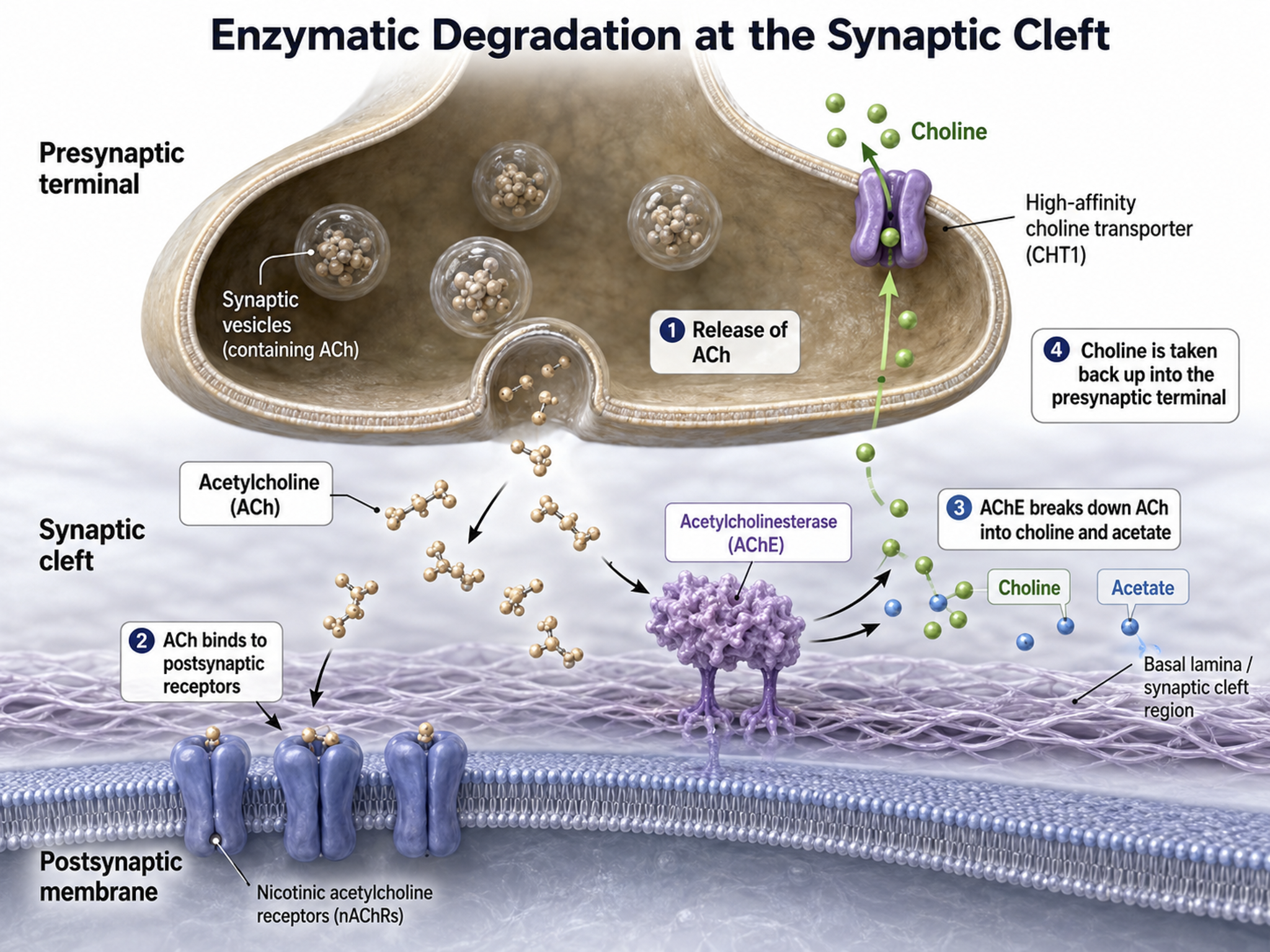
In enzymatic degradation, enzymes located in the synaptic cleft and the cytoplasm of the presynaptic neuron's terminal button split neurotransmitter molecules apart (e.g., acetylcholine).
Electrical Synapses
Electrical synapses communicate information across gap junctions between adjacent membranes using ions. Gap junctions are narrow spaces between two cells bridged by connexons (protein channels) that allow ions near-instantaneous travel. Gap junction illustration adapted from © VectorMine/Shutterstock.com.
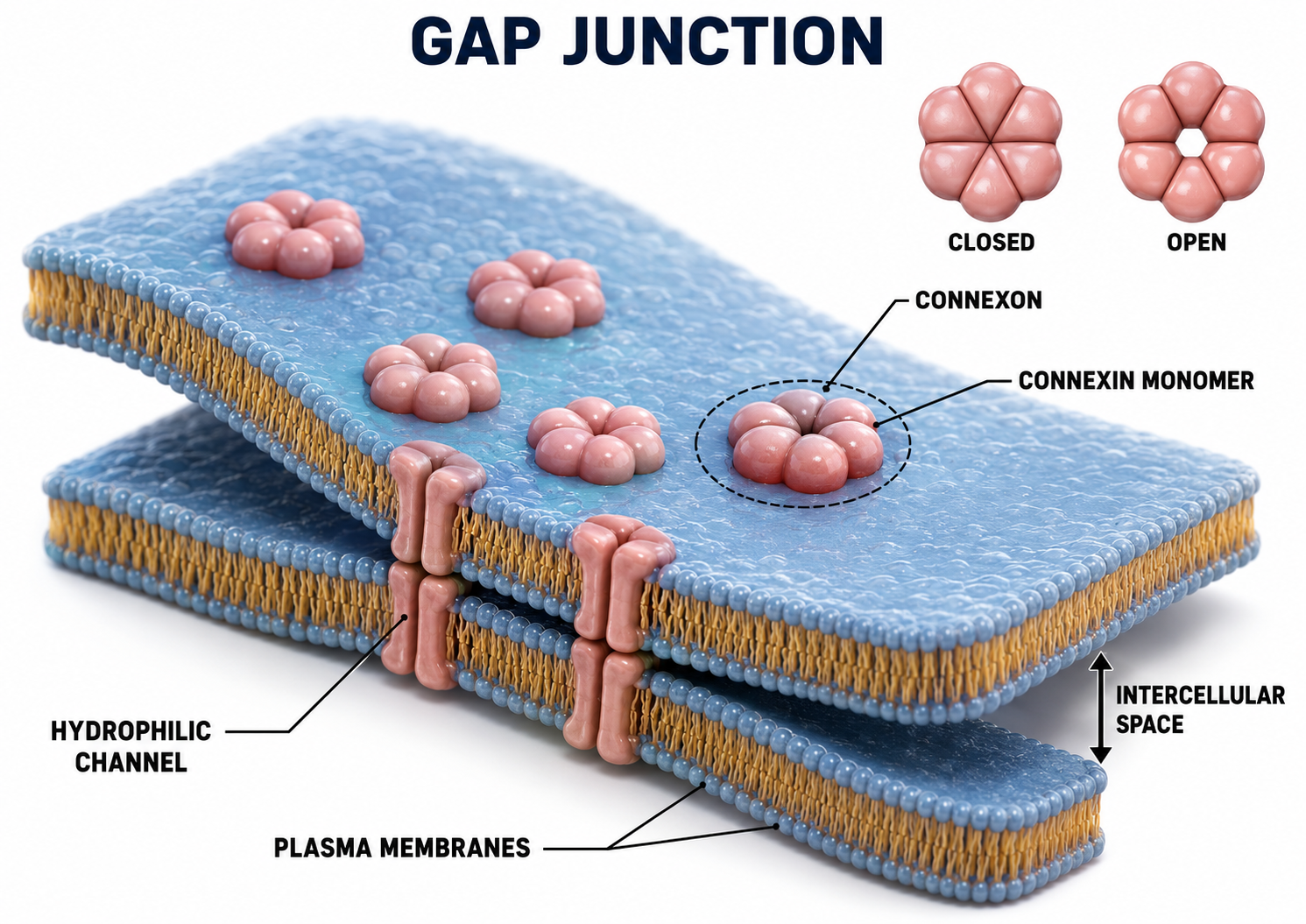
Electrical synapses are generally symmetrical. Ions flow across a 3-nm gap junction into the more negatively charged neuron as long as the gap junction remains open, meaning that whether neurons are presynaptic or postsynaptic depends on their respective charges. When two neurons are electrically coupled, an action potential in one induces a postsynaptic potential (PSP) in the paired neuron.
Transmission across electrical synapses is nearly instantaneous, compared with the roughly 0.5-1-ms synaptic delay at chemical synapses. The rapid information transmission that characterizes electrical synapses enables large circuits of neurons to synchronize their activity and fire simultaneously, a property that is directly relevant to the synchronized oscillations observed in the EEG.
Studies in recent years have revealed that electrical synapses are common in every part of the mammalian CNS. When two neurons are electrically coupled, an action potential in the presynaptic neuron causes a small amount of ionic current to flow across the gap junction channels into the other neuron. This current causes an electrically mediated postsynaptic potential (PSP) in the second neuron. Note that, because most electrical synapses are bidirectional, when that second neuron generates an action potential, it will in turn induce a PSP in the first neuron (Bear et al., 2020, p. 113).
Neurons that secrete hormones use electrical synapses to release their chemical messengers simultaneously. Neonatal brains may use gap junctions to activate many neurons at once. Image of long, fibrous astrocyte processes using Golgi's silver chromate technique © Jose Luis Calvo/Shutterstock.com.
Gap junctions may be a preliminary step toward developing chemical synapses between these neurons, eventually replacing their electrical synapses. Prenatally and postnatally, gap junctions enable nearby neurons to coordinate their development by sharing electrical and chemical communications (Bear, Connors, & Paradiso, 2026; Breedlove & Watson, 2023).
Old school view: synapses are either electrical or chemical.
New school view: synapses can be both electrical and chemical.
Key Takeaways
Neurons communicate chiefly through chemical synapses, releasing neurotransmitters from vesicles by exocytosis across the synaptic cleft to bind ionotropic receptors for fast effects or metabotropic receptors for slower, longer-lasting ones. Communication is not limited to the cleft, because volume transmission, axonal varicosities, and retrograde signaling let neurotransmitters influence broader regions. The nervous system fine-tunes these signals through modulation of both transmitter release and receptor sensitivity, acting more like a volume control than an on-off switch. More than two hundred neurotransmitters have been identified, and a given transmitter can be excitatory or inhibitory depending on the receptor it activates. Electrical synapses complement chemical ones by allowing near-instantaneous, synchronized firing across coupled neurons.
Check Your Understanding
- How do ionotropic and metabotropic receptors differ in their speed and duration of action, and why does that distinction matter for the EEG?
- What are three ways neurons release neurotransmitters outside of a classical synapse, and why is this significant?
- How do presynaptic facilitation and presynaptic inhibition change synaptic communication without altering the action potential itself?
- Why can the same neurotransmitter produce opposite effects at different synapses?
- How does transmission across electrical synapses differ from transmission across chemical synapses?
Discoveries Since Graduate School
This section highlights major neuroscience discoveries that have reshaped our understanding of neural communication since many clinicians completed their training. These advances, from adult neurogenesis to the mirror neuron system, have direct implications for how we conceptualize and deliver neurofeedback interventions.
Neuroscientists have learned a great deal more about neuron-to-neuron communication since graduate school. The most important findings are that the adult brain creates new neurons, silent synapses may mediate neuroplasticity in adulthood, the lymphatic system extends to the brain, neuronal networks exhibit mirroring properties, and neurons can release more than one NT, release NTs outside of a synapse, conduct two-way conversations, modulate NT release and action, talk to astrocytes that enclose synapses, and electrically communicate almost instantaneously.
Neurogenesis
Neuroscience has challenged the long-held doctrine that the adult human brain does not create new neurons. Neurogenesis, the creation of new neurons in adults, was first demonstrated in the human hippocampus by Eriksson et al. (1998), and recent evidence favors its persistence there, but the question is still actively contested rather than settled. Adult neurogenesis in the olfactory bulb is well established in rodents (Lim & Alvarez-Buylla, 2016) but is not supported by carbon dating of human tissue. Neurogenesis elsewhere in the adult human brain remains more controversial still. Animal research has yielded evidence of functionally significant neurogenesis in the amygdala, caudate nucleus and putamen (striatum), cortex, hypothalamus, and substantia nigra (Jurkowski et al., 2020). The neurogenesis graphic by Rebeca Cuesta is licensed under the Creative Commons Attribution-Share Alike 4.0 International license.
Silent Synapses
Silent synapses are inactive due to the absence of glutamate AMPA receptors. Researchers studying adult mice discovered these synapses on the ends of threadlike filopodia, thin, exploratory projections that extend from neurons. The simultaneous firing of two neurons connected by a silent synapse causes missing AMPA receptors to appear on the filopodia cell membrane and remodel it to resemble a dendritic spine (Vardalaki et al., 2022).
The next step is to determine whether the adult human brain also contains silent synapses. If it does, they represent a potential target for increasing cognitive flexibility in the elderly and may help explain how neurofeedback training can produce changes even in mature brains. Filopodia photomicrograph by Aurea D. Sousa and Richard E. Cheney under the Creative Commons Attribution-Share Alike 4.0 International license.
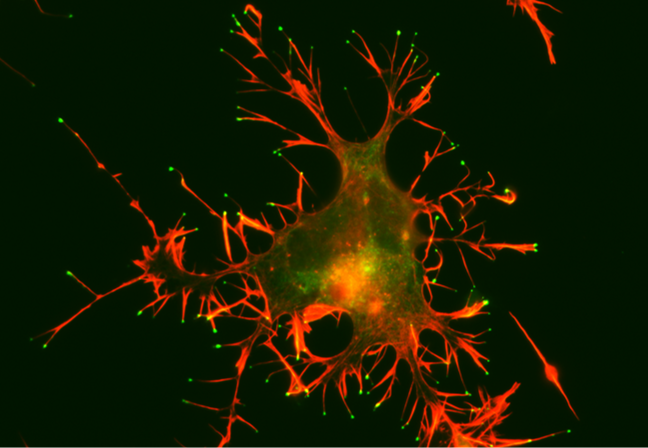
A CAD cell (a neuronal cell line) expressing GFP-Myo10 (green) was stained for actin filaments (red) to visualize the slender cellular protrusions known as filopodia. Overexpressing Myo10 induces large numbers of filopodia and is responsible for the unusually large number of filopodia on this cell.
Holly Barker (2022) writing for The Scientist, explained:
The study may explain how the brain is able to learn new things without having to sacrifice existing connections, the researchers say. The ability of the brain to use different synapses 'solves the plasticity versus flexibility dilemma,' says Harnett. If all the brain's synapses are flexible, then you can't preserve old information. But if they're all stable, then it is difficult to learn new things, he says. Instead, the brain employs both: spiny synapses for stability and filopodia for flexibility.But instead of distinct categories, Harnett's group is beginning to think about dendritic projections as existing on a continuum, from filopodia on one end to mature spines at the other. 'It is a spectrum of maturity, strength, and plasticity,' says study author Dimitra Vardalaki, a PhD candidate in Harnett's lab.
Cutting Edge
The discovery of silent synapses in adult mice is a very recent development (Vardalaki et al., 2022). Because these dormant synapses sit on filopodia and can be activated without disturbing established connections, they may let the brain learn new information while preserving old memories. If the adult human brain contains them as well, they offer a plausible cellular route by which neurofeedback could reshape mature circuits. Confirming their presence in humans is an active and important research question.
Mirror Neuron System
Researchers discovered primate neurons with both motor and visual properties in the premotor cortex. The mirror properties are due to a neuron's connections and not its construction. The cortex graphic adapted from © Vasilisa Tsoy/Shutterstock.com.
These mirror neurons fired when primates grasped and manipulated objects, and when another primate or human performed the same action (di Pellegrino et al., 1992; Rizzolatti & Craighero, 2004). Mirroring extends across species, including facial expressions.
Molenberghs et al. (2011) unexpectedly found neurons with mirroring properties in the cerebellum, limbic system, and primary visual cortex. Graphic adapted from Wikimedia Commons.
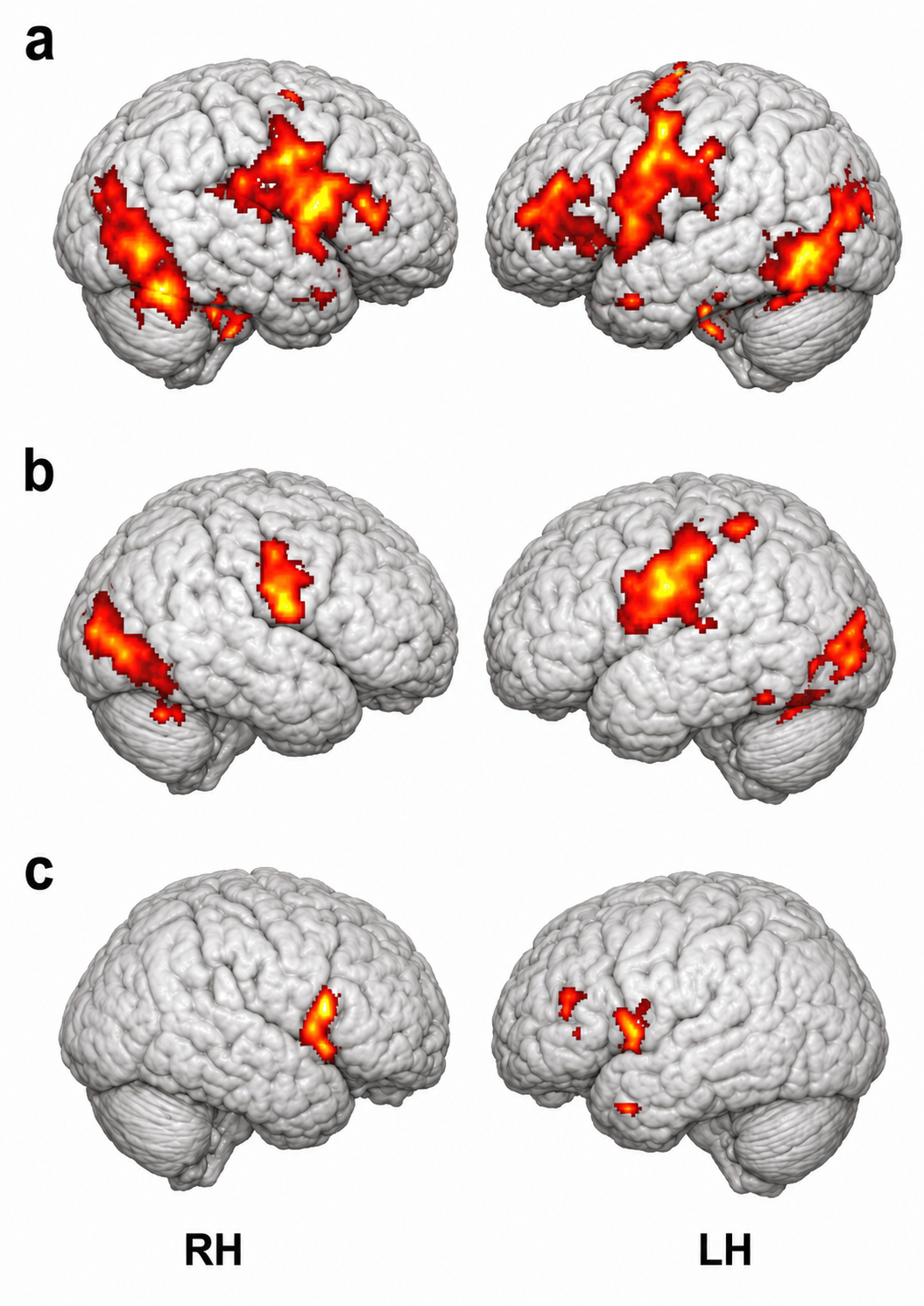
The authors proposed that a core network is responsible for observing and executing movements. The nervous system recruits additional areas to perform non-motor affective, auditory, and somatosensory functions.
Mirror neurons look like other neurons when examined using a microscope. Their mirror properties emerge from their connections within sensory, motor, and emotional systems. Perhaps most mirror neurons may be tuned by experience (Catmur, Walsh, & Heyes, 2007).
The mirror neuron system (MNS) appears to encode the goal of a motor act and its component movements, whether a model manipulates an object or mimes the action. The MNS encodes the actions of others and stores them to predict their future actions (Rajmohan & Mohandas, 2007).
Soon after birth, an immature mirror neuron system may allow babies to imitate their parents' mouth movements, like thrusting out the tongue.

Ramachandran (2011) has called the mirror neurons activated when they observe others' movements "monkey see, monkey do neurons." He calls mirror neurons activated by others' emotional displays "Gandhi neurons." Check out Ramachandran's TED Talk, The Neurons that Shaped Civilization.
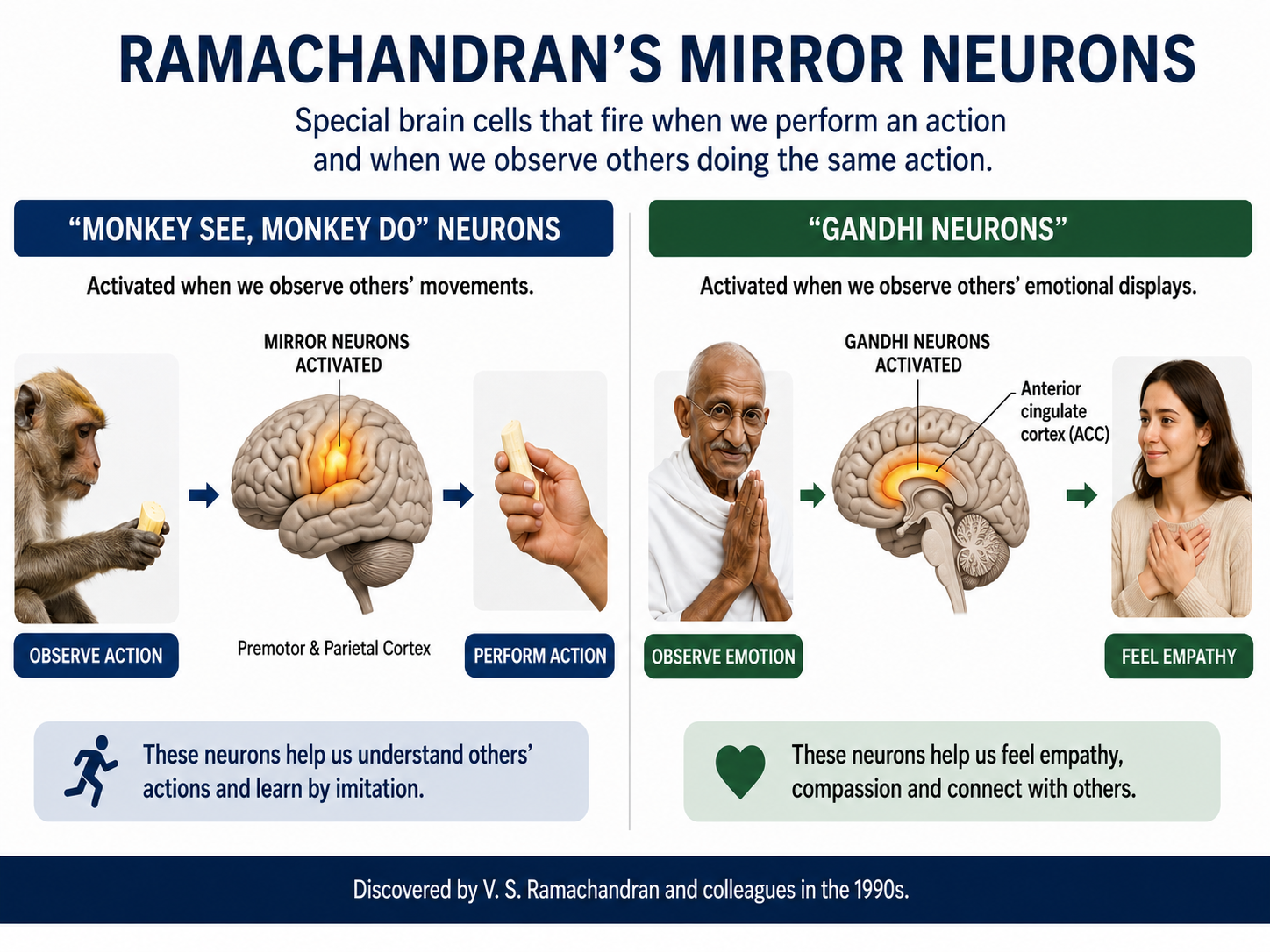
Investigators have speculated that the human MNS may mediate empathy, imitation learning, language, social cognition, and theory of mind (Buccino et al., 2006; Rajmohan & Mohandas, 2007; Schmidt et al., 2021).
Rizzolatti and Sinigaglia (2008) hypothesized that the primary role of the MNS is to help us understand others' intentions, which allows us to achieve empathy. When we observe others' facial expressions of emotion, visual information may be directly transmitted to mirror neurons in the insula, producing the visceral changes that color our emotions.
In autism, mirror neurons may not fire when observing other individuals performing actions. This may help explain deficits in empathy, social skills, language, and the development of a theory of mind (Enticott et al., 2012).
Heyes et al. (2021) summarized the current state of our knowledge about the MNS.
For action understanding, multivoxel pattern analysis, patient studies, and brain stimulation suggest that mirror-neuron brain areas contribute to low-level processing of observed actions (e.g., distinguishing types of grip) but not to high-level action interpretation (e.g., inferring actors' intentions). In the area of speech perception, although it remains unclear whether mirror neurons play a specific, causal role in speech perception, there is compelling evidence for the involvement of the motor system in the discrimination of speech in perceptually noisy conditions. For imitation, there is strong evidence from patient, brain-stimulation, and brain-imaging studies that mirror-neuron brain areas play a causal role in copying of body movement topography. In the area of autism, studies using behavioral and neurological measures have tried and failed to find evidence supporting the 'broken-mirror theory' of autism.
Cortical Architecture
This section describes the organization of the cerebral cortex, including its gray and white matter, convolutions, and layered structure. Understanding cortical architecture is essential because the arrangement and orientation of cortical neurons determines what the scalp EEG can and cannot detect.
While no one has counted the neurons in the human nervous system directly, the best current estimate is that an adult brain contains about 86 billion neurons (Azevedo et al., 2009; see also Voytek, 2013). As noted earlier, cortical neurons average several thousand synapses each; the brain-wide average is lower, because most neurons are small cerebellar granule cells with only a few inputs. The once-standard claim that glia outnumber neurons ten to one has been retracted. Isotropic-fractionator counts give a glia-to-neuron ratio below 1:1 and fewer than 100 billion glial cells in the human brain (Azevedo et al., 2009; von Bartheld et al., 2016). Glial somas are considerably smaller than neuronal somas, roughly 6 to 10 μm in diameter (Hammond, 1996), although a single astrocyte's processes span a far larger territory. Animation © nmlfd/iStockphoto.com.
The cerebral cortex comprises neuronal cell bodies, glial cells, and blood vessels. Beneath the neocortex lie myelinated nerves (white matter), unmyelinated fibers, and glial cells.
The cerebral cortex covers the cerebral hemispheres and consists of gray and white matter. Gray (or grey) matter, which looks grayish brown, comprises cell bodies. White matter gains its opaque white color from myelinated axons. The cerebral cortex is shown below.
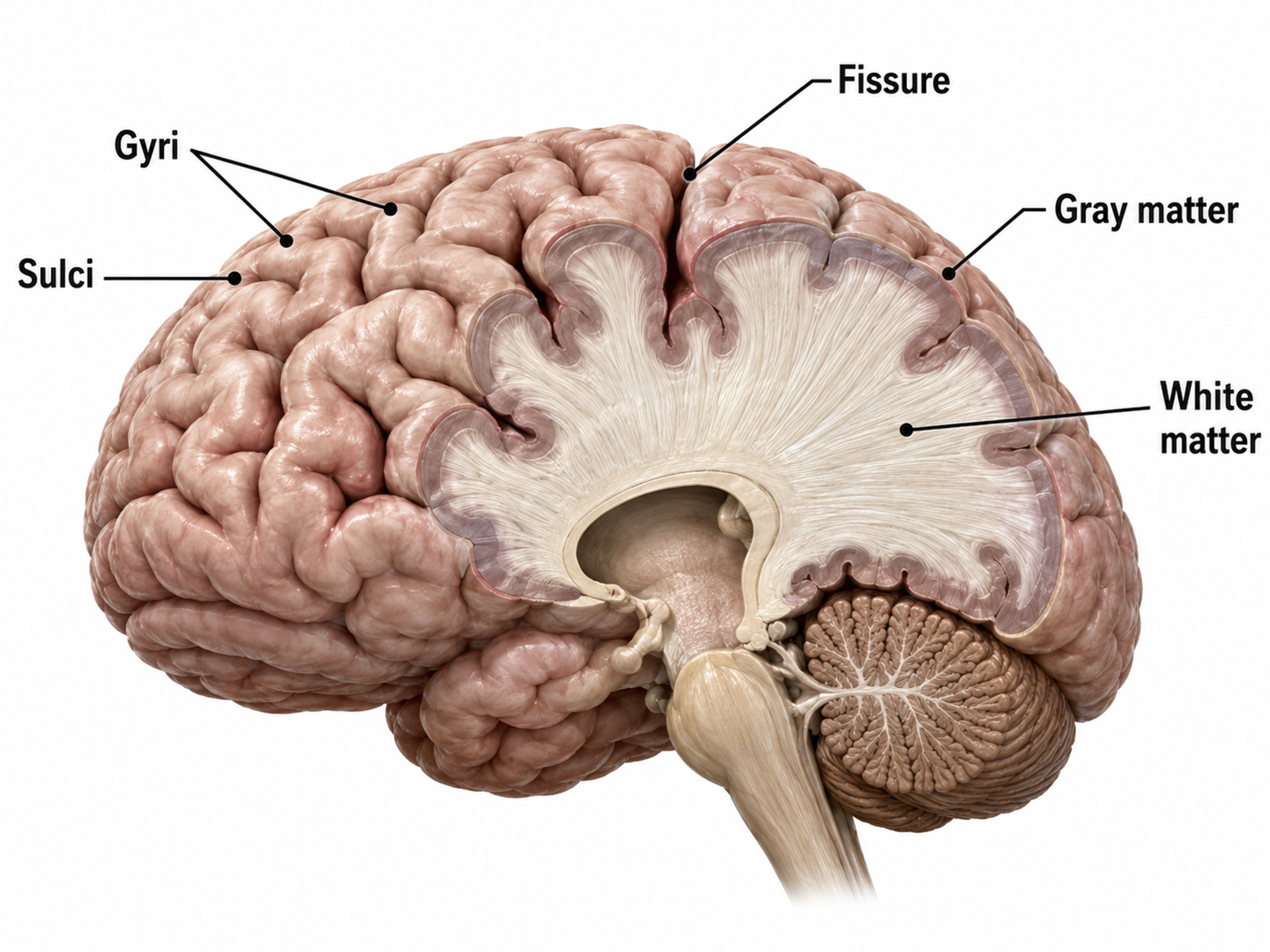
The convolutions of the cerebral cortex bury about two-thirds of its surface area within the sulci and maximize the amount of cortical tissue housed within the skull. Cerebral cortical convolutions include sulci, which are shallow grooves in the surface of the cerebral hemisphere (central sulcus), fissures, which are deep grooves (lateral fissure), and gyri, which are ridges of cortex demarcated by sulci or fissures (precentral gyrus) (Carlson & Birkett, 2021).
There are two main types of cortex: neocortex and allocortex.
The neocortex or isocortex consists of six layers, 1 to 4.5 mm thick and averaging about 2.5 mm, with a total surface area usually given as roughly 2,000 to 2,500 cm2 and white matter underneath. Layers I-III receive corticocortical afferent fibers that connect the left and right hemispheres. Layer III is the main source of corticocortical efferent fibers.
Layer IV is the primary destination of thalamocortical afferents and intra-hemispheric corticocortical afferents. Layer V is the primary origin of efferent fibers that target subcortical structures that have motor functions. Layer VI projects corticothalamic efferent fibers to the thalamus, which together with the thalamocortical afferents, creates a dynamic and reciprocal relationship between these two structures (Creutzfeldt, 1995).
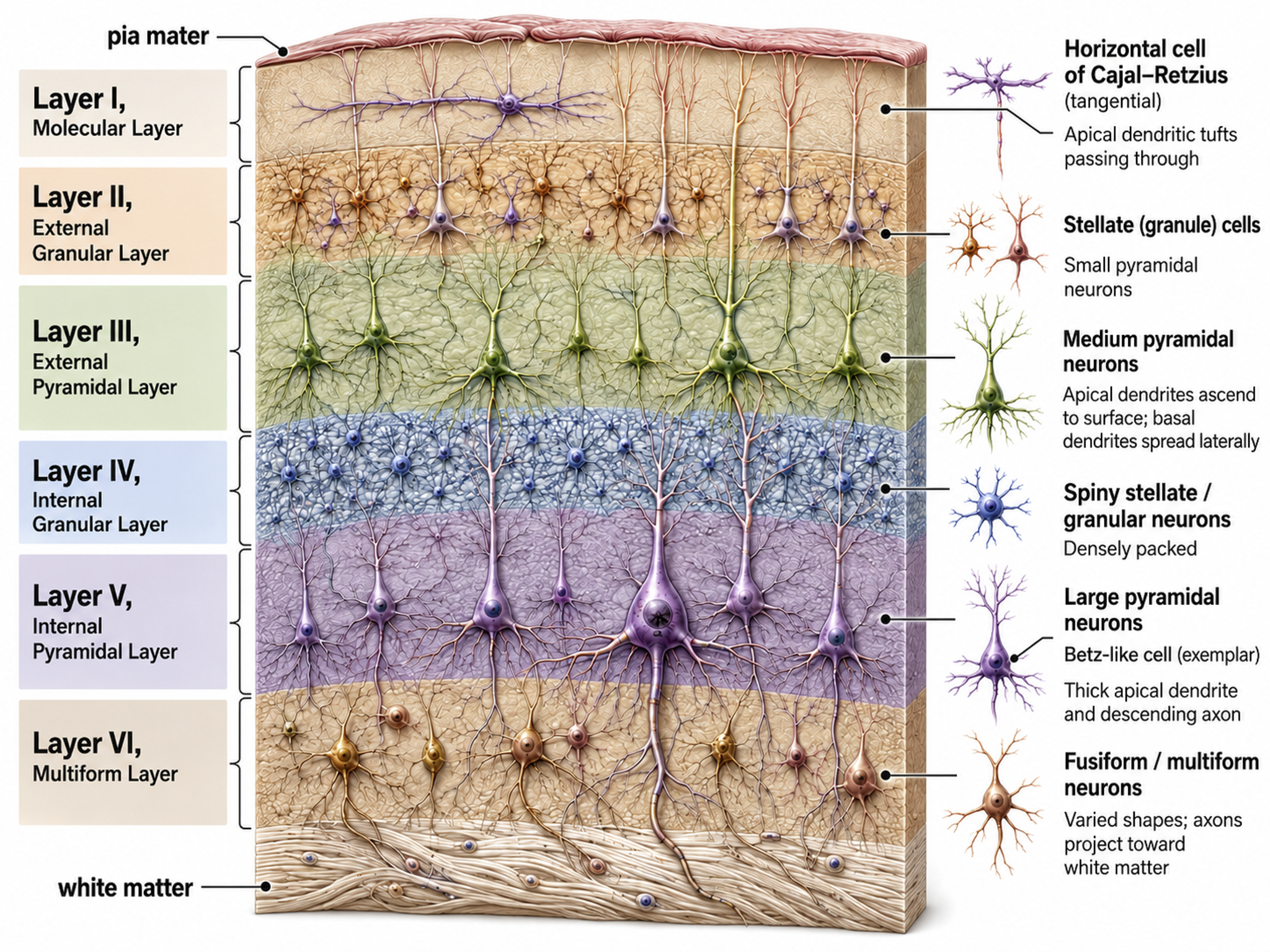
Allocortex, which means other cortex, usually has between three or four layers, compared with the neocortex's six layers. The allocortex has less volume than the neocortex and comprises the olfactory system and hippocampus.
A transitional region between the neocortex and allocortex is called the paralimbic cortex.
For a basic overview of the cortex, watch the Khan Academy video Cerebral Cortex.
Neurons in the Cortex
We can classify cerebral cortical neurons by whether their dendrites display spines. Spiny neurons, which have either pyramidal or stellate (star-like)-shaped cell bodies, are usually excitatory. While all pyramidal cells are spiny neurons, stellate cells can be spiny or aspinous (Bear, Connors, & Paradiso, 2026). Pyramidal neurons are especially important for neurofeedback because their parallel alignment and perpendicular orientation to the cortical surface make them the primary generators of the EEG signal detected at the scalp.
The graphic below shows spiny and aspinous dendrites.
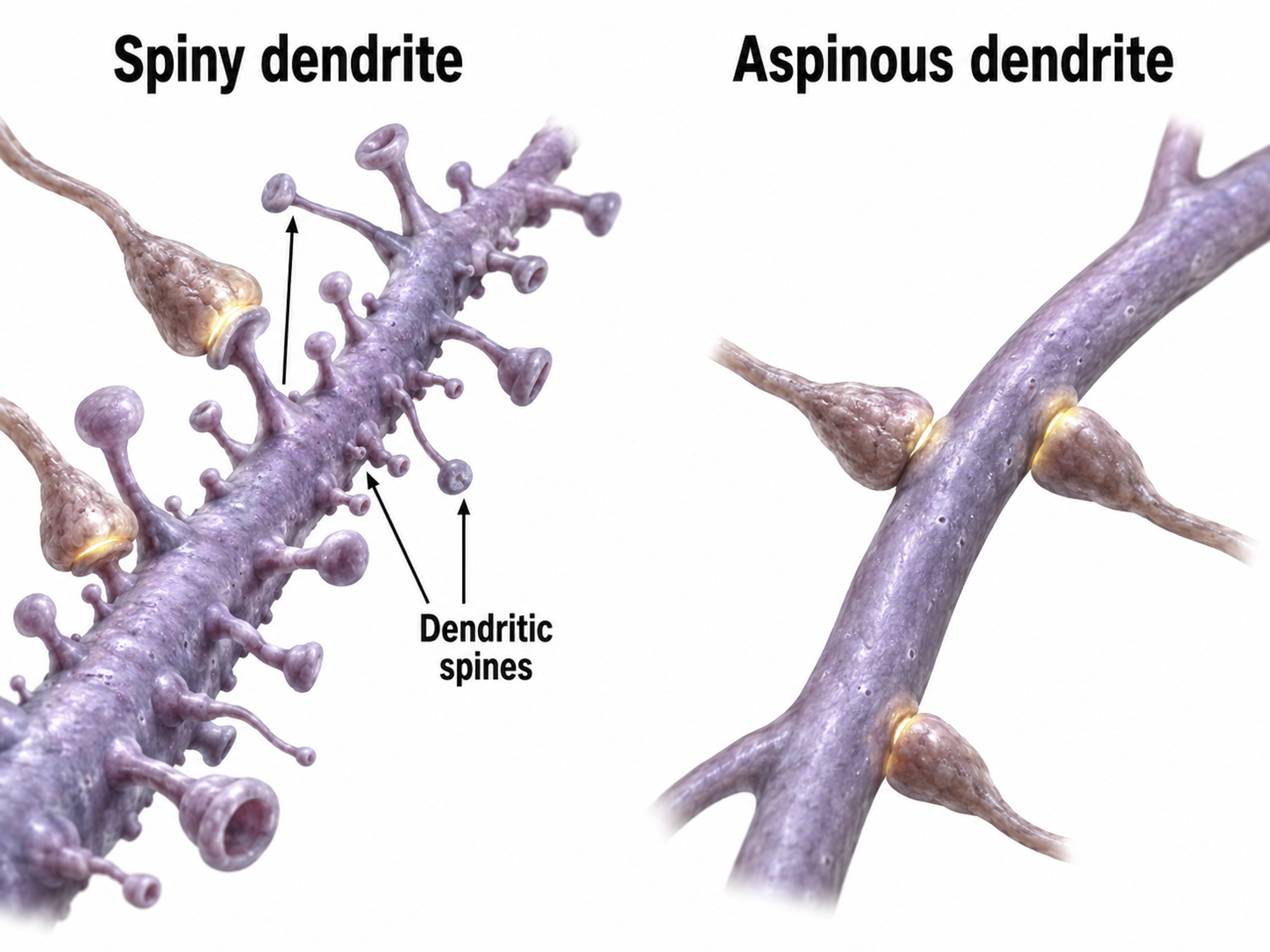
Dr. John C. Fiala, and Dr. Kristen M. Harris created this reconstruction of a dendritic spine. Creative Commons Attribution-Share Alike 3.0.
_and_massive_aggregation_of_polyribosomes_(black)..jpg)
There are many types of aspinous (smooth) neurons which are believed to be inhibitory.
What is the EEG?
This section explains what the scalp EEG actually measures, how it is generated by pyramidal neurons in cortical columns, and the roles of local field potentials, dipole generators, amplitude, and frequency. This knowledge is foundational for every neurofeedback clinician because it connects the electrical activity displayed on a neurofeedback screen to the underlying neural processes being trained.
The scalp EEG is the voltage difference between two recording sites recorded over time. The EEG is primarily generated by large pyramidal neurons located in layers 3 and 5 of the cortical gray matter, which ranges from about 1 to 4.5 mm thick and averages about 2.5 mm. Image of a pyramidal neuron revealed using Golgi silver chromate © Jose Luis Calvo/Shutterstock.com. Note that the cell body's apical dendrite, which projects toward the cortical surface, and basilar dendrites, which arise from the base, feature an extensive network of spines.
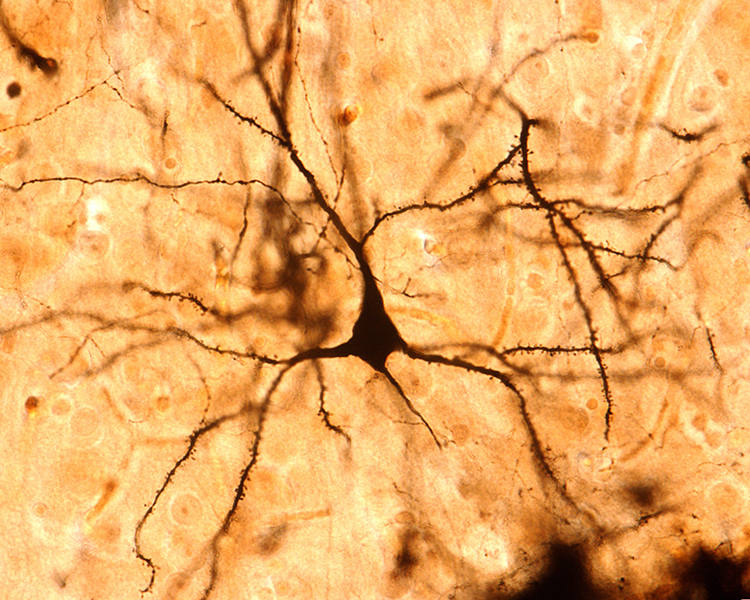
Local activity is a composite of local and network influences. Network communication systems and local cortical functions show different characteristics across the cortex and produce unique and specific EEG patterns in different regions.
The movie below is a BioTrace+/NeXus-32 display of the raw EEG with voltage shown as μV peak to peak © John S. Anderson.
What Can the EEG Tell Us?
With the EEG, we can follow the progression from stimulus to behavioral response. This allows us to determine whether each step in the processing chain is functioning correctly and identify causal factors in dysfunctional outcomes. For clinicians, this capability makes the EEG an invaluable tool for both assessment and treatment planning.
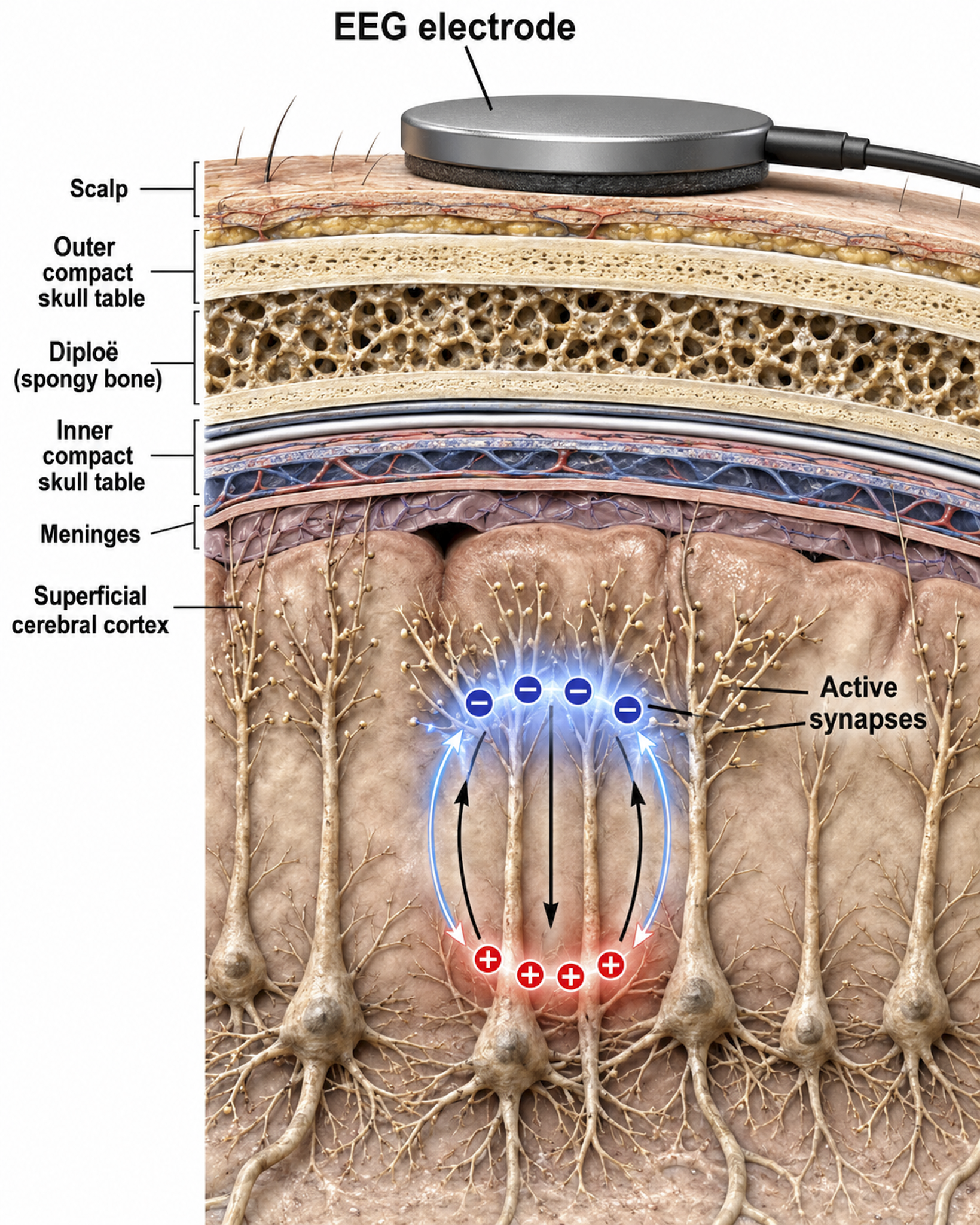
Source of the Scalp EEG
The scalp EEG results from the summation of large areas of gray matter activity. These areas are polarized synchronously due to the input of oscillatory or transient evoked activity, and they comprise thousands of cortical columns containing large pyramidal cells aligned perpendicularly to the cortical surface.
Pyramidal neurons of the cerebral cortex stained with the Golgi silver chromate © Jose Louis Calvo/Shutterstock.com.
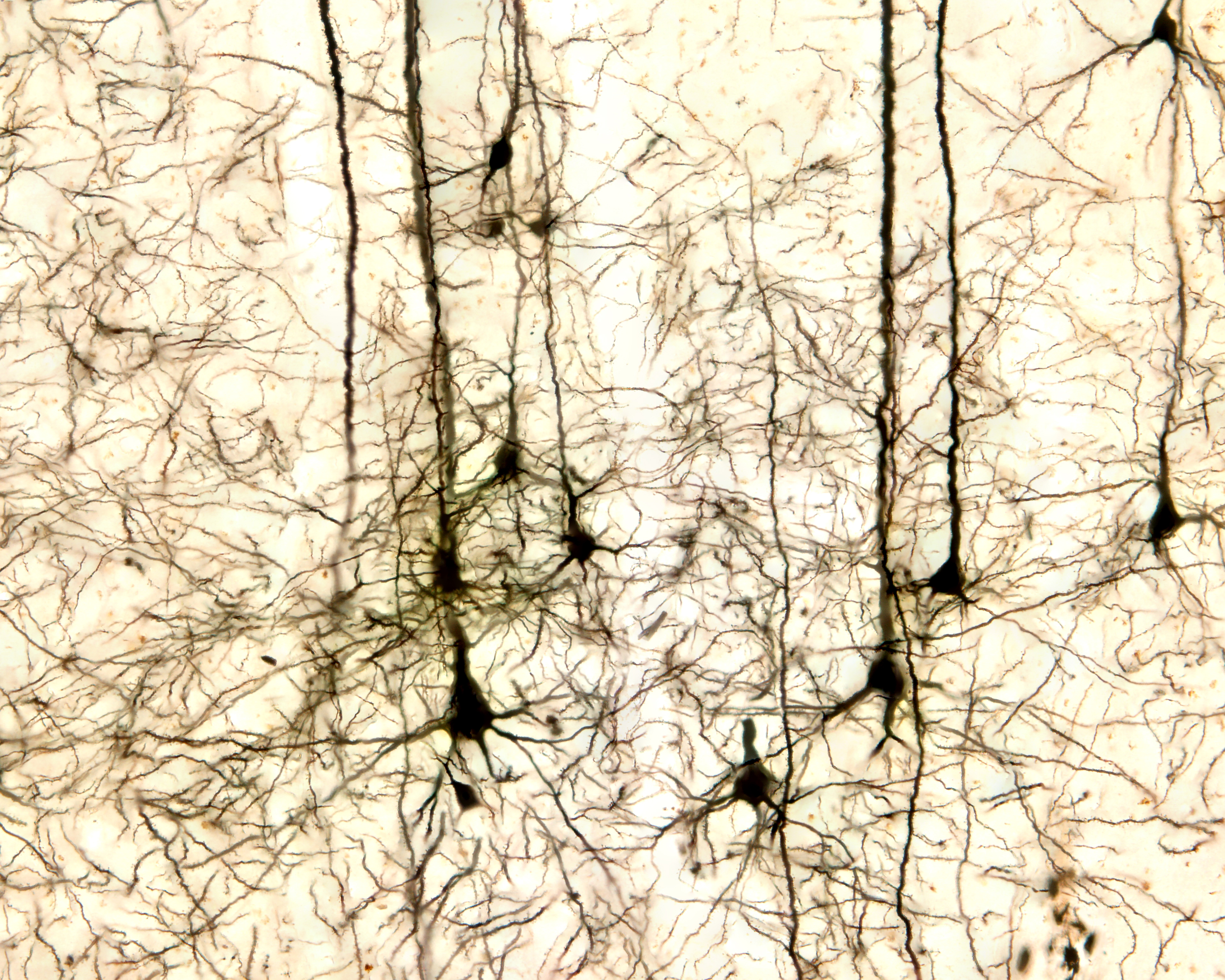
Pyramidal neurons are found in all cortical layers except layer 1 and represent the primary type of output neuron in the cerebral cortex. Pyramidal neuron graphic © Kateryna Kon/Shutterstock.com.
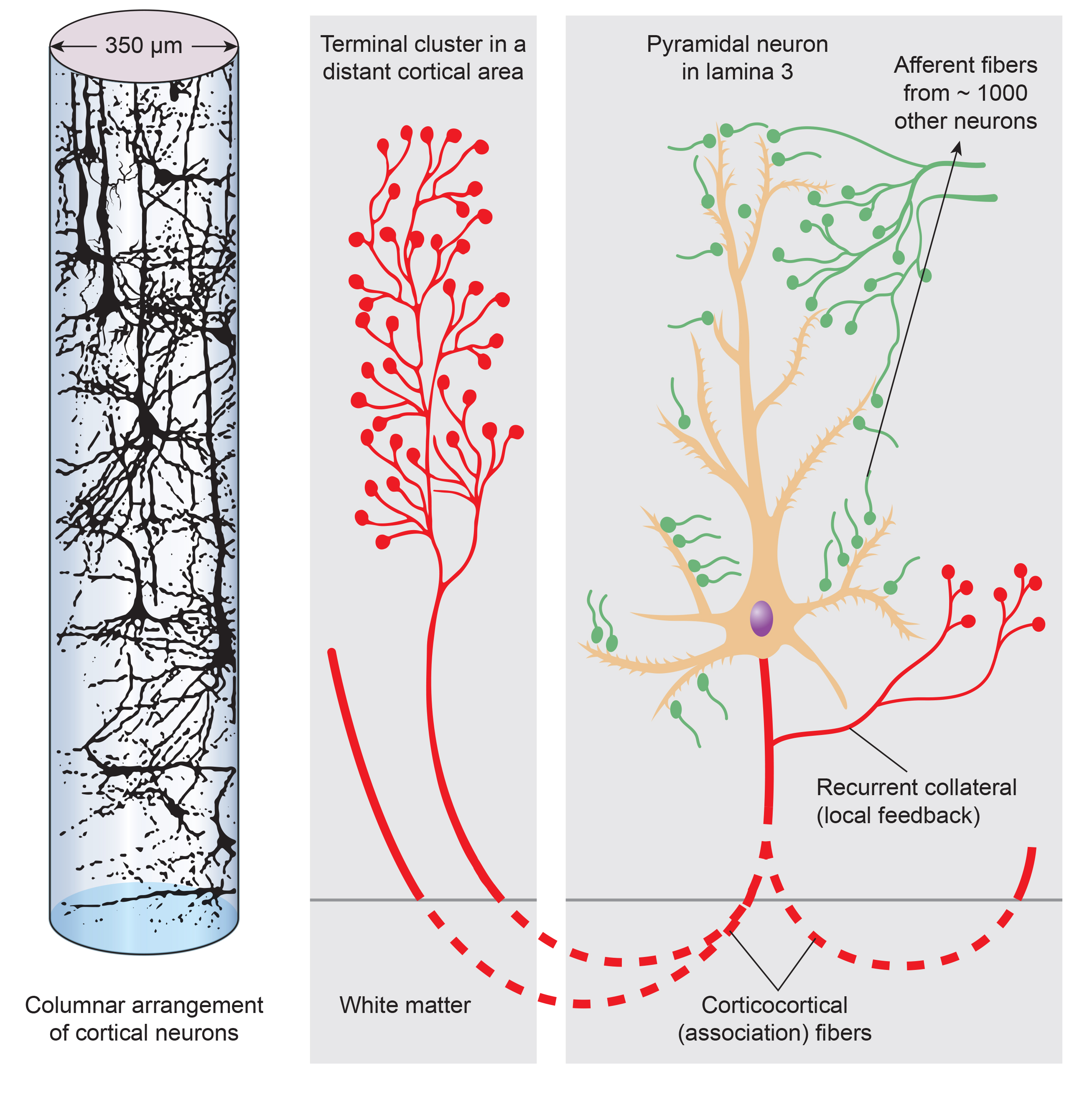
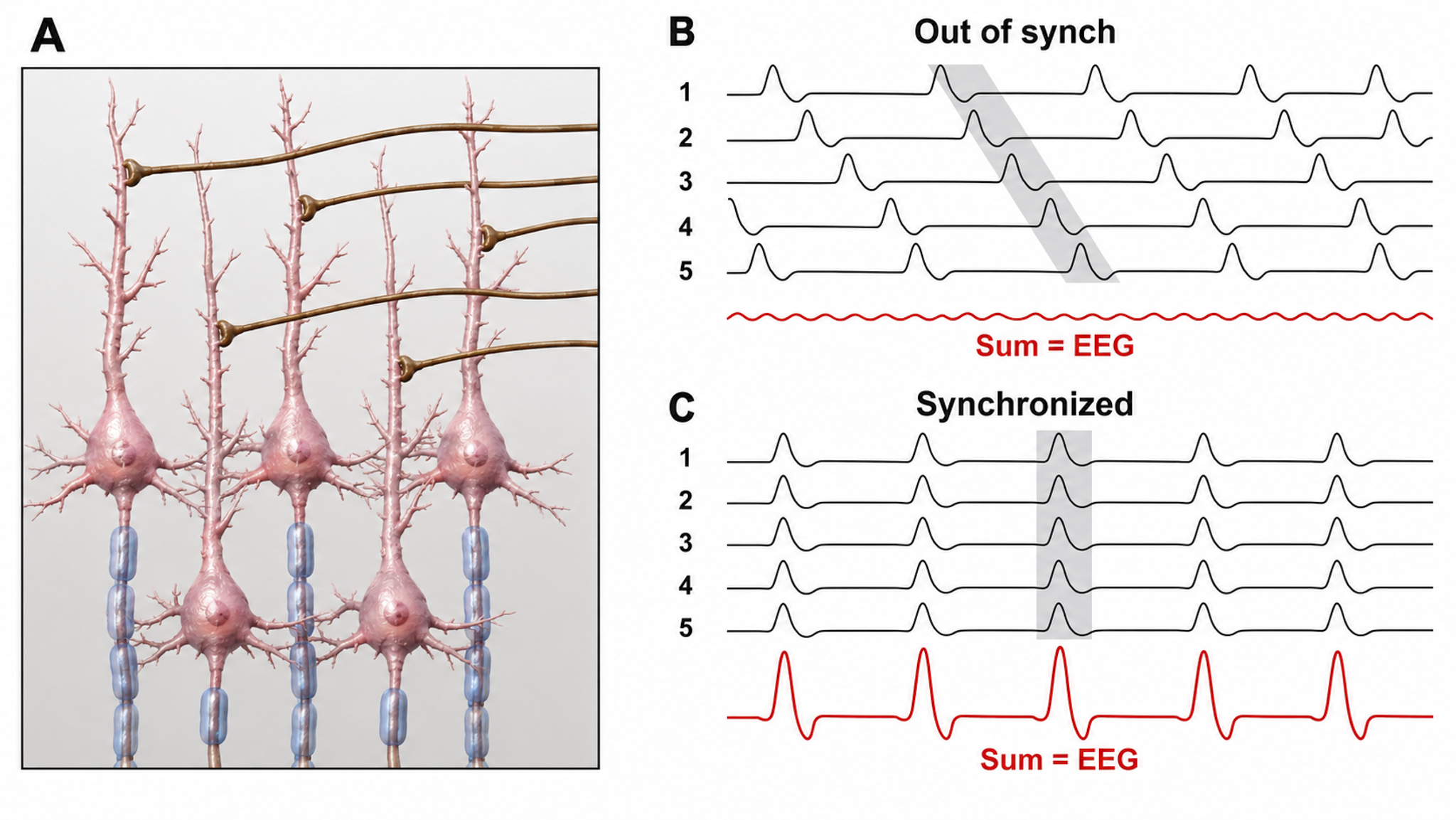
The scalp EEG results from the summation of EPSPs and IPSPs in thousands of cortical columns containing large pyramidal cells perpendicular to the cortical surface. The columns are synchronously polarized (made more negative) and depolarized (made less negative) due to the input of oscillatory or transient evoked activity. Graphic redrawn by minaanandag on Fiverr.com.
Artist: Dani S@unclebelang. This WEBTOON is part of our Real Genius series.

Local Field Potentials
The local field potential (LFP) is the aggregate electrical effect of interconnected pyramidal neurons firing within cortical columns, plus additional mechanisms like glial cell modulation of the cortical electrical gradient. The LFP is the bridge between individual neuron activity and the macroscopic EEG signal; it reflects the collective behavior of local neural populations.
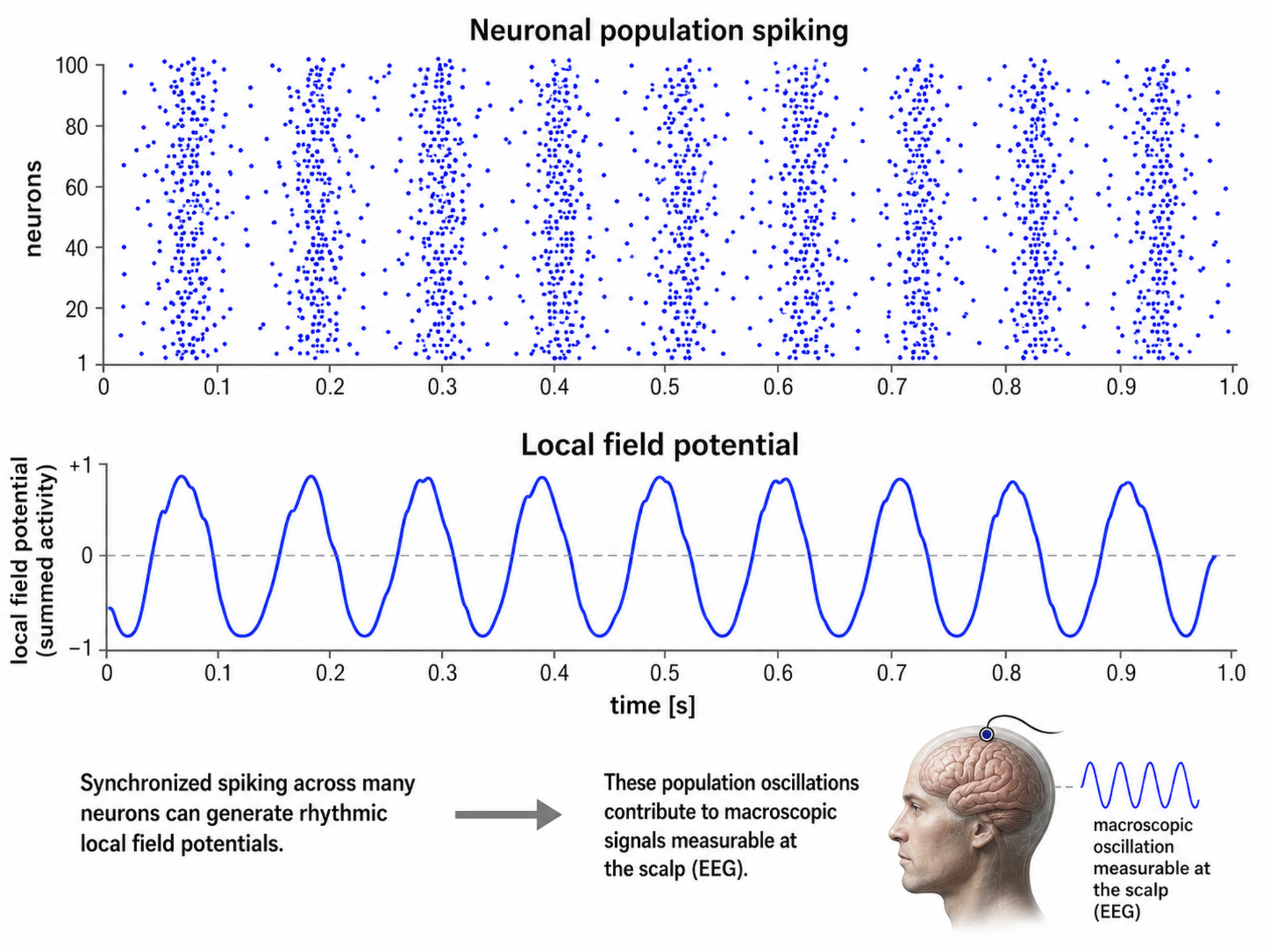
Caption from Wikipedia's article on Neural Oscillation. Simulation of neural oscillations at 10 Hz. The upper panel shows spiking of individual neurons (with each dot representing an individual action potential within the population of neurons). On the lower panel, the local field potential reflects their summed activity. This figure illustrates how synchronized patterns of action potentials may result in macroscopic oscillations that can be measured outside the scalp.
Do not confuse the "spiking" of individual neurons with epileptogenic spikes in the scalp EEG.
Scalp Electrical Potentials
Scalp electrical potentials represent the sum of all available electrical fields. Fields of opposite polarity (+/-) cancel each other out so that scalp potentials are greater when large aggregates of neurons polarize and depolarize synchronously. The scalp EEG represents a weighted sum of all active currents within the brain that generate open fields, including non-cortical sources.
Action potentials reflect neuronal output. In extracellular recordings they appear as brief events lasting less than 2 ms, isolated by high-pass filtering above roughly 300 Hz. The familiar 100-mV swing of an action potential is an intracellular measurement; recorded extracellularly, a spike is only tens to a few hundred microvolts. Action potentials play a minor role in the scalp EEG because their amplitude falls below 60 µV outside a 50-μm (0.050-mm) radius of the cell body. Since scalp electrodes are several centimeters from cortical neurons and are generally aligned away from the scalp, action potentials are unlikely to contribute significant voltages to the scalp EEG. This is a key point: what we measure on the scalp during neurofeedback are postsynaptic potentials, not action potentials.
Local Field Potentials Regulate Neuron Excitability and Firing
Neurons are most likely to fire during the depolarizing phase of the local field potential. Neurons are more excitable when they are "in phase" with the LFP and are inhibited when they are out of phase. Thus, at any instant of time, the amplitude and frequency of the EEG are regulated by the LFP, which in turn is influenced by oscillatory mechanisms such as slow cortical potentials. This rhythmic gating of neural excitability is what makes neurofeedback training possible; by modifying these oscillatory patterns, clients can influence which neural populations fire and when.
The movie is a 19-channel BioTrace+ /NeXus-32 display of SCPs © John S. Anderson. Negative SCPs drift up, and positive SCPs drift down, depending on the software settings. However, the convention in electroencephalography is to show negative up. SCPs represent a global shift in DC voltage across the cortex and reflect a generally higher (negative SCPs) or lower (positive SCPs) state of cortical excitability regulating neural networks.
The EEG is a moment-to-moment measure of the excitability of action potential firing, like gates opening and closing on the half cycle. The synchronous activity of large pyramidal neurons networked in cortical columns creates the EEG.
The Composition of the EEG
The EEG is composed of electrical potentials, varying in two dimensions: frequency and amplitude.
Sources of IPSP and EPSP Inputs
Many sources contribute input that results in IPSP and EPSP activity within cortical neurons. These sources primarily contribute influences such as oscillatory generator input or ascending event-related evoked input.
EEG Sources
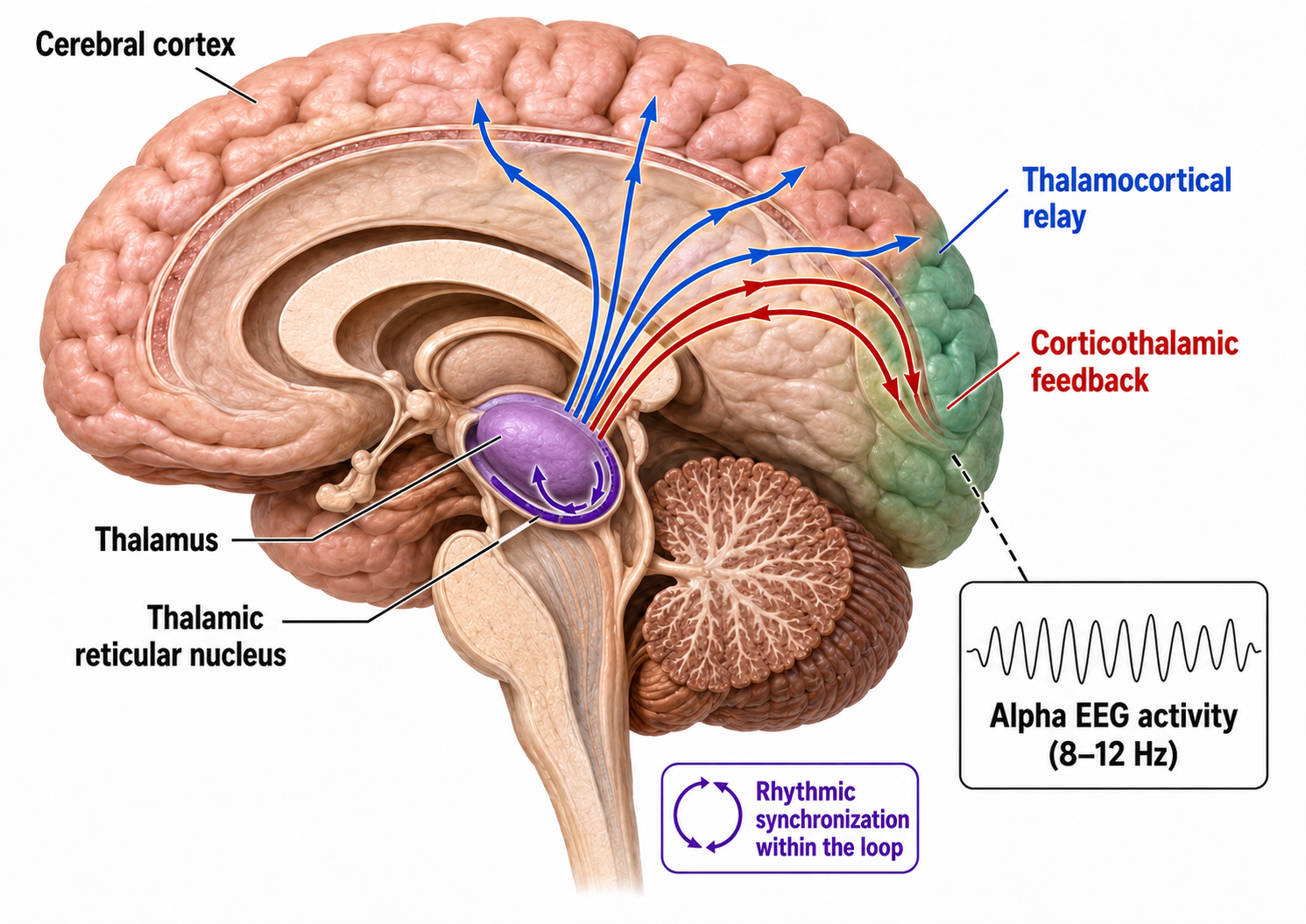
Generators like the thalamus produce oscillatory activity among many interconnected neurons, including EEG patterns like the alpha rhythm. The thalamus functions as the brain's central relay station, and its rhythmic output shapes much of the EEG activity that clinicians observe during recording.
Movie © John S. Anderson. The recording begins with eyes open. The eyes-closed condition starts at 14'01" and clearly shows increased 8-12 Hz voltage (posterior dominant rhythm or PDR) in occipital and parietal locations in the line tracing and topographic maps to the right of the tracing.
The eyes open again at 14'31", and alpha attenuates (alpha blocking). This demonstrates the posterior dominant rhythm (generally known as "alpha") appearing in the eyes-closed condition when visual sensory input is stopped, and the attenuation or blocking of this rhythm as sensory input returns in the eyes-open condition. This eyes-open/eyes-closed comparison is one of the most common assessment procedures in clinical neurofeedback.
The thalamus contributes to slow cortical potentials, 1-4 Hz delta, 8-12 Hz alpha, and beta activity in the 20-38 Hz range, as well as to 40-Hz gamma activity. The diagram shows the connections between the pulvinar (bottom right) and reticular nuclei (bottom left) of the thalamus and the cortex © Elsevier Inc. - Netterimages.com.
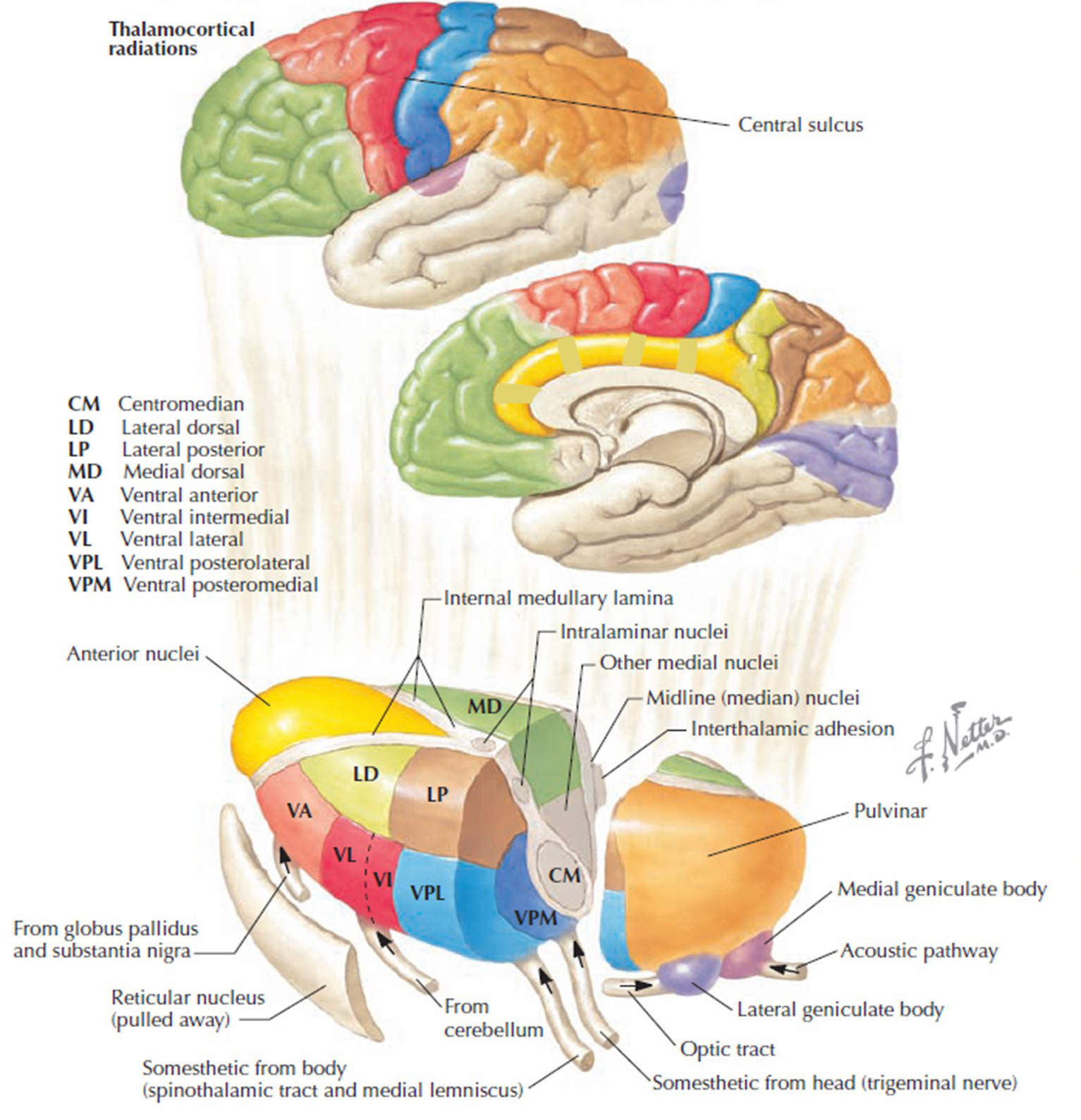
Thalamocortical (signaling from the thalamus to the cortex) cells are subject to excitatory drive from their system afferents, from monosynaptic corticothalamic (signaling from the cortex to the thalamus) fibers, and from the brainstem reticular formation (ascending reticular activating system, ARAS). They receive inhibitory drive from local interneurons and neurons in the reticular nucleus of the thalamus (RNT).
Note that the RNT neurons are excited by activity in thalamocortical cells and corticothalamic cells. The connections are precisely organized; each column in a primary cortical area sends corticothalamic fibers back to the same part of its specific thalamic nucleus that sends its thalamocortical fibers to that cortical column. The corticothalamic fibers also synapse on the RNT cells receiving input from that part of the thalamic nucleus. Each cortical receiving area is said to be "reciprocally connected" with its specific thalamic nucleus. Like the thalamocortical cells, RNT cells and cortical neurons also receive excitatory drive from the ARAS (Jackson & Stoney, 2006).
The EEG is generated by thalamocortical (delta and alpha), medial cortical and subcortical regions (theta), and cortico-cortical (beta) sources.
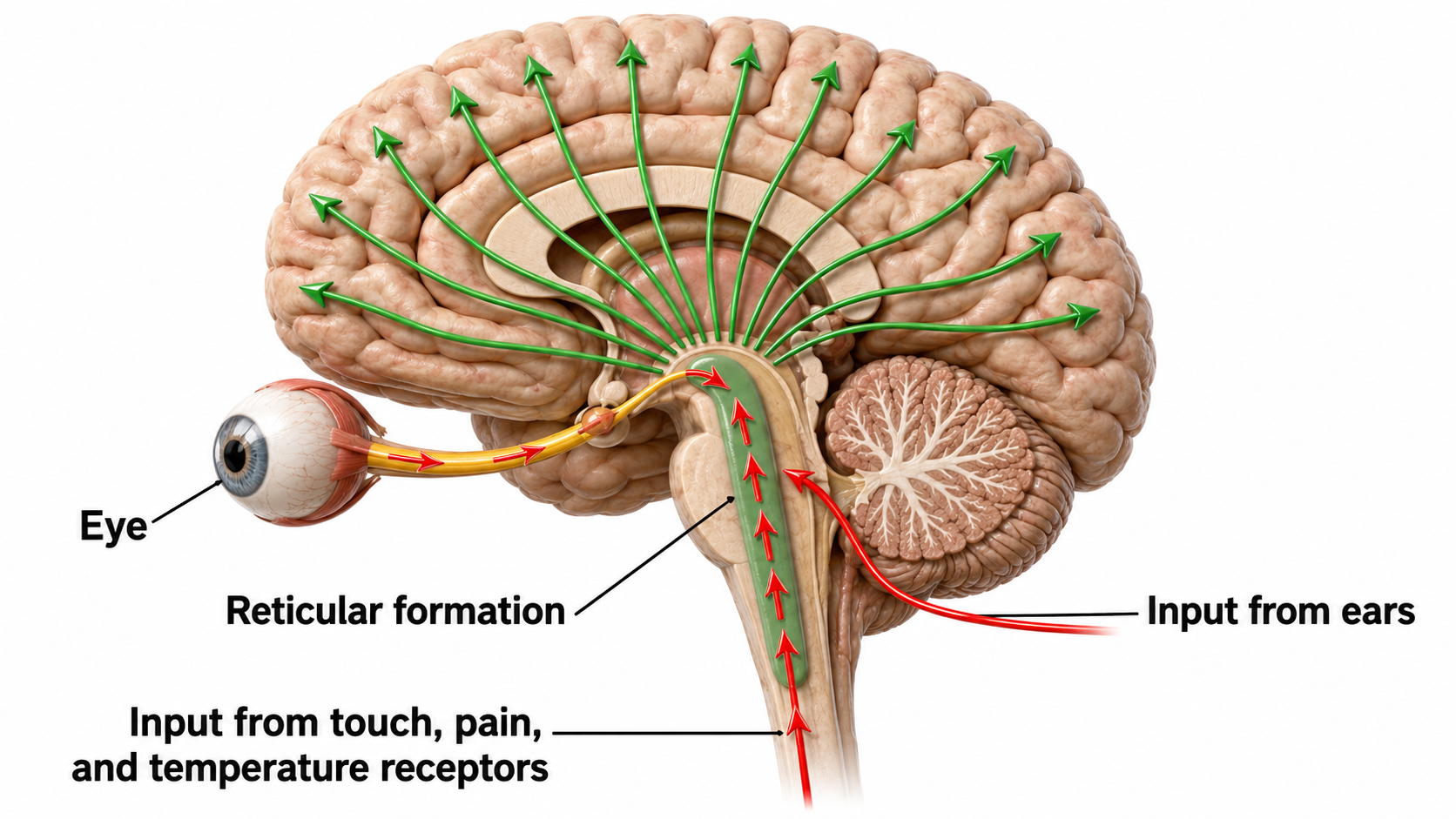
Ascending reticular activating system input helps drive the cortical responses recorded as event-related potentials following diverse stimuli like a flashing light or a sound. Event-related potentials (ERPs) are the brain's response to externally applied stimuli, events, or cognitive/motor tasks. They are time-locked measures of brain electrical activity.
Dipole Generators
Large cortical pyramidal neurons organized in macrocolumns are oriented with an apical dendrite projecting toward the scalp and an axon descending in the opposite direction. An "Equivalent Dipole Generator" usually represents the sum of all multipolar current sources. Summed generators are modeled as dipoles to aid the conceptual understanding of the electrical fields involved. Graphic adapted from Lipping (2017).
EEG Signals (Brainwaves)
The EEG represents changes in a brain area's electrical activity (potential) compared to a "neutral" site or another brain area. The EEG is displayed as oscillations or voltage fluctuations, which show a "wave" pattern when plotted on a graph.
"These oscillations are generated spontaneously in several areas of the cerebral cortex as neuronal networks transiently form assemblies of synchronously firing cells." Klaus Linkenkaer-Hansen.
Sink, Source, and Dipole
We can describe pyramidal cells in terms of their sink, source, and dipole. A sink (-ve), which may be located at the bottom, middle, or top of the apical dendrite, is where positive ions enter the dendrite. Cation (positive ion) entry gives the extracellular space a negative charge. The source (+ve) is where the current exits the cell. Finally, the dipole is the field created between the sink and source (Thompson & Thompson, 2015).
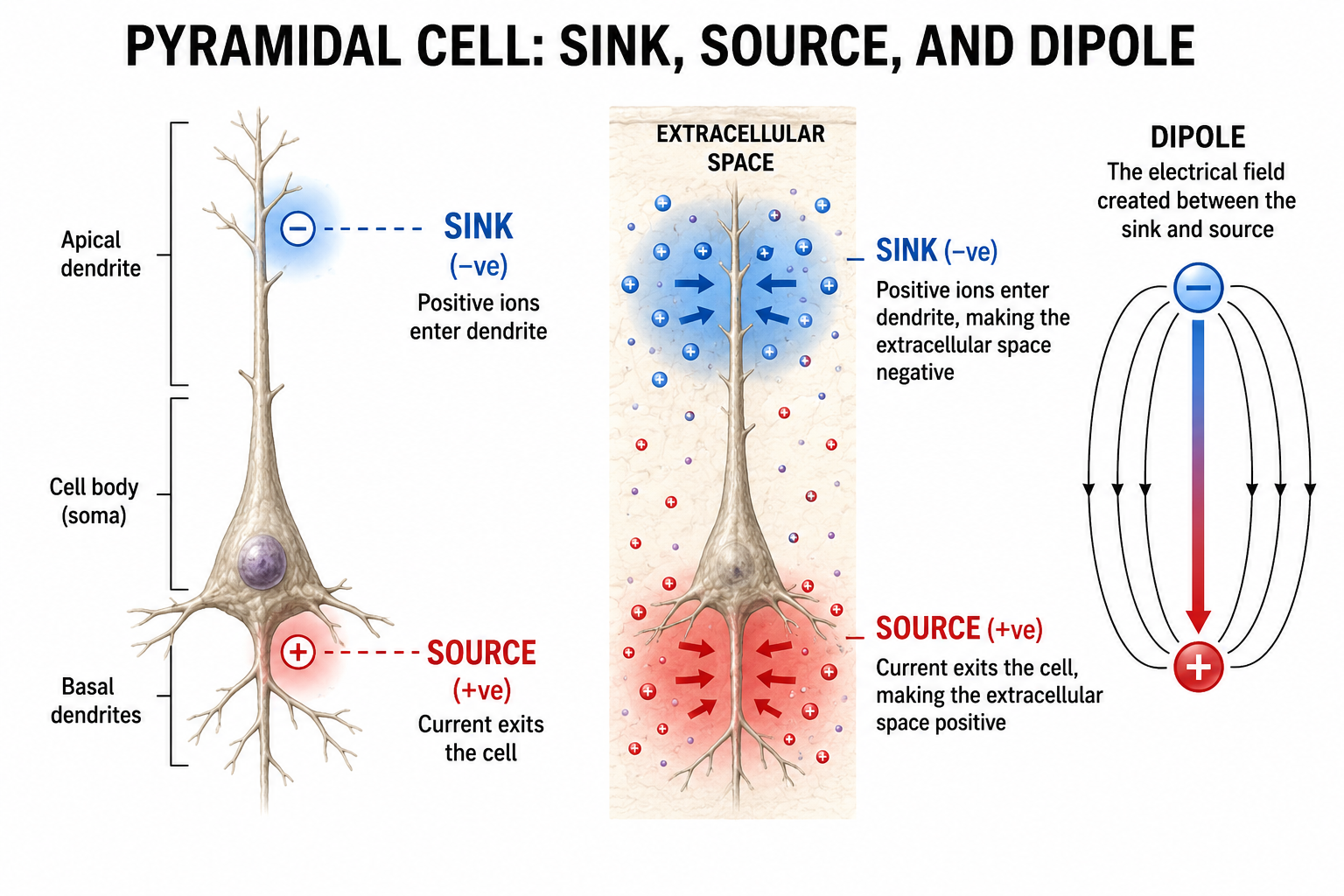
The postsynaptic potentials (EPSPs and IPSPs) propagated by the apical dendrites in layers 2 and 3 create an extracellular dipole layer parallel to the cortical surface. The dipole layer's electrical polarity is the opposite of the deeper cortical layers 4 and 5 (Fisch, 1999).
A cortical dipole is created when pyramidal neurons depolarize simultaneously, a phenomenon called local synchrony. Synchrony matters more than sheer numbers. Because asynchronously active neurons contribute potentials that partly cancel, a comparatively small synchronized population can dominate the recorded signal; roughly 6 to 10 cm2 of contiguous, synchronously active cortex is generally taken as the minimum needed to register at the scalp. A small fraction of these neurons firing in step can produce visible changes in EEG feedback. This creates the potential for operant conditioning to help clients learn to modify EEG activity through neurofeedback.
Cortical dipoles have three properties: site (depends on source), size (oscillation frequency and voltage), and relative position with respect to sulci and gyri (Collura, 2014).
The EEG is Mainly Sensitive to Radially Oriented Dipoles
Evolution has convoluted the human brain to increase its computing power without enlarging the skull. This enfolding has created two easily visible anatomical features: gyri and sulci.
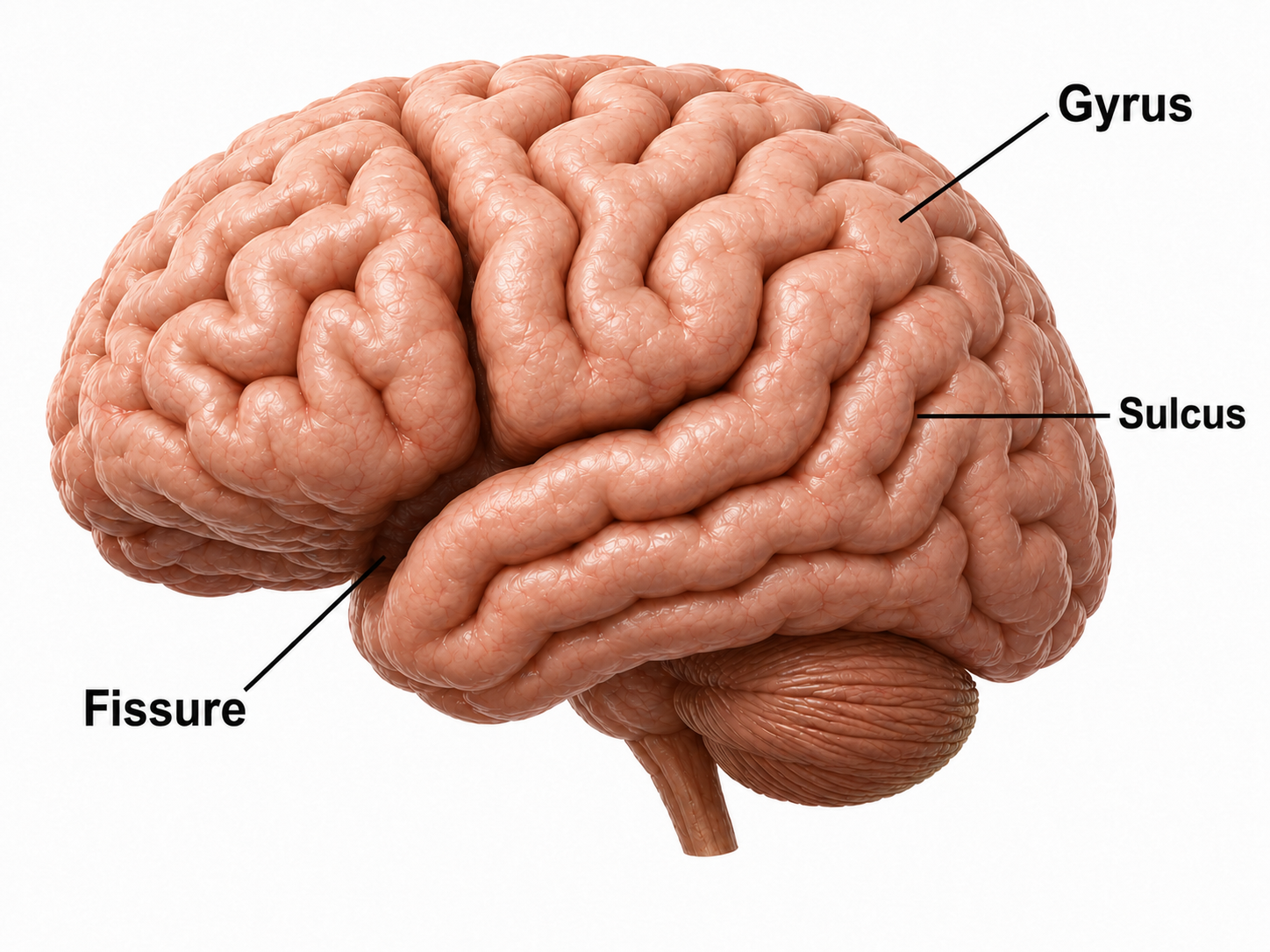
Recall that a gyrus is a ridge of the convoluted cerebral cortex, while a sulcus is a valley.
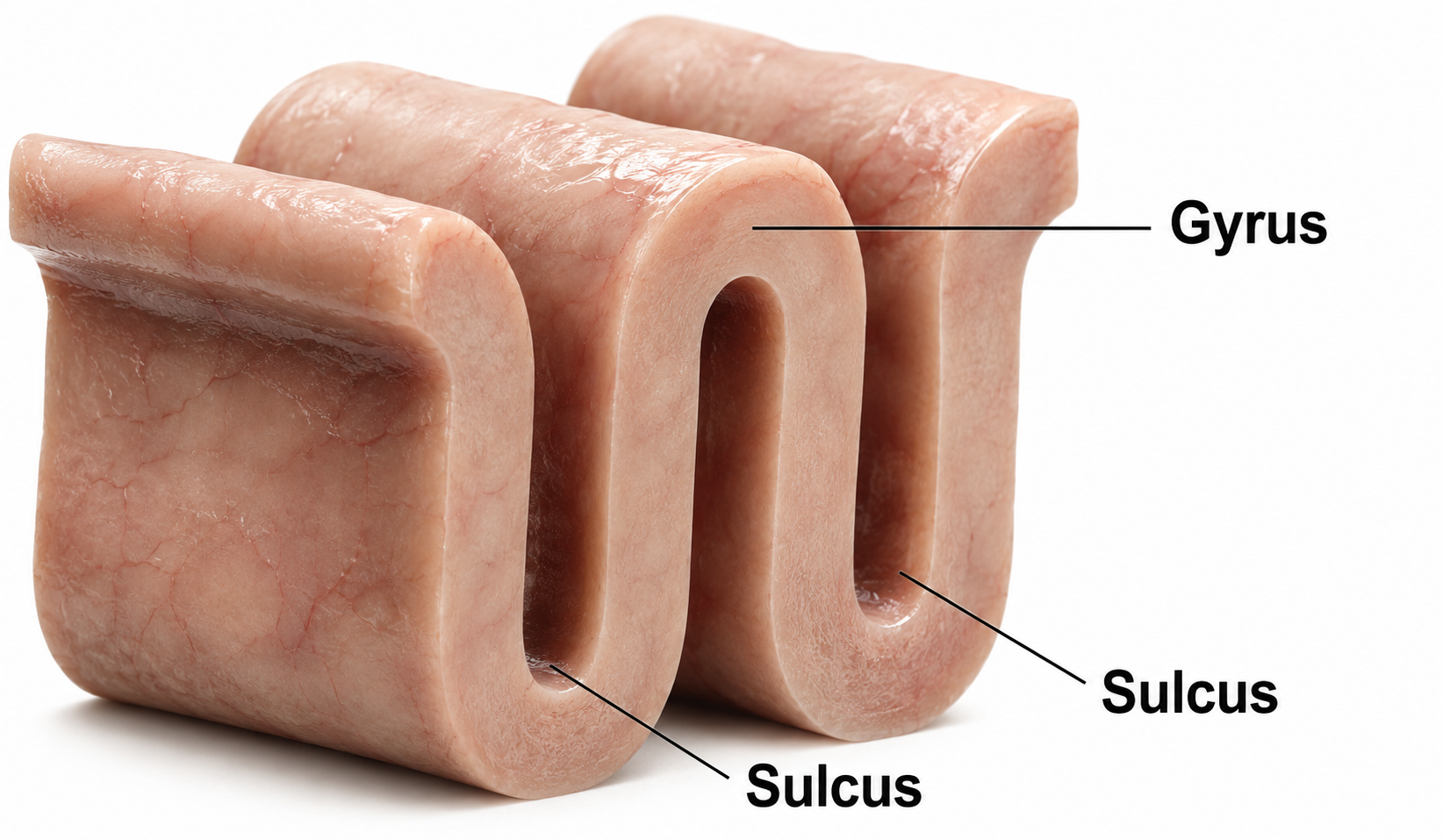
A dipole's orientation relative to the scalp determines how well the EEG detects it. A radial dipole points outward from a gyral crown, roughly perpendicular to the overlying scalp, so its field projects directly toward the recording electrodes and is detected well. A tangential dipole lies in the wall of a sulcus, oriented parallel to the scalp, so much of its field runs sideways beneath the electrodes rather than toward them. When the two facing walls of a sulcus generate opposing tangential dipoles, their fields largely cancel at the surface. This geometry is why the scalp EEG favors activity from gyral crowns and can underrepresent sources buried in sulci (Bear, Connors, & Paradiso, 2026).
The EEG is most sensitive to a correlated dipole layer in gyri. The EEG is less sensitive to a correlated dipole layer in sulci, valleys within the cortex. Finally, the EEG is insensitive to an opposing dipole layer in sulci. This sensitivity pattern is a key limitation of scalp EEG that neurofeedback clinicians should keep in mind: the EEG preferentially detects activity from gyral surfaces and may miss activity originating deep within sulci.
The signal also spreads as it travels from cortex to scalp, a process called volume conduction. The brain, cerebrospinal fluid, skull, and scalp act as a passive conductor that smears each cortical source across a broad area of the scalp, so a single electrode records a blend of contributions from many underlying generators. Volume conduction is the main reason scalp EEG has limited spatial resolution and why an electrode's reading depends heavily on the choice of reference. It should not be confused with volume transmission, the chemical process described earlier (Collura, 2014).
The EEG is composed of electrical potentials that vary along the dimensions of amplitude and frequency.
EEG Amplitude
The "amount" of voltage, or amplitude reflects the number of neurons firing in synchrony at a given frequency. Lower neuron firing rates and fewer neurons firing in synchrony correspond to lower signal amplitude.
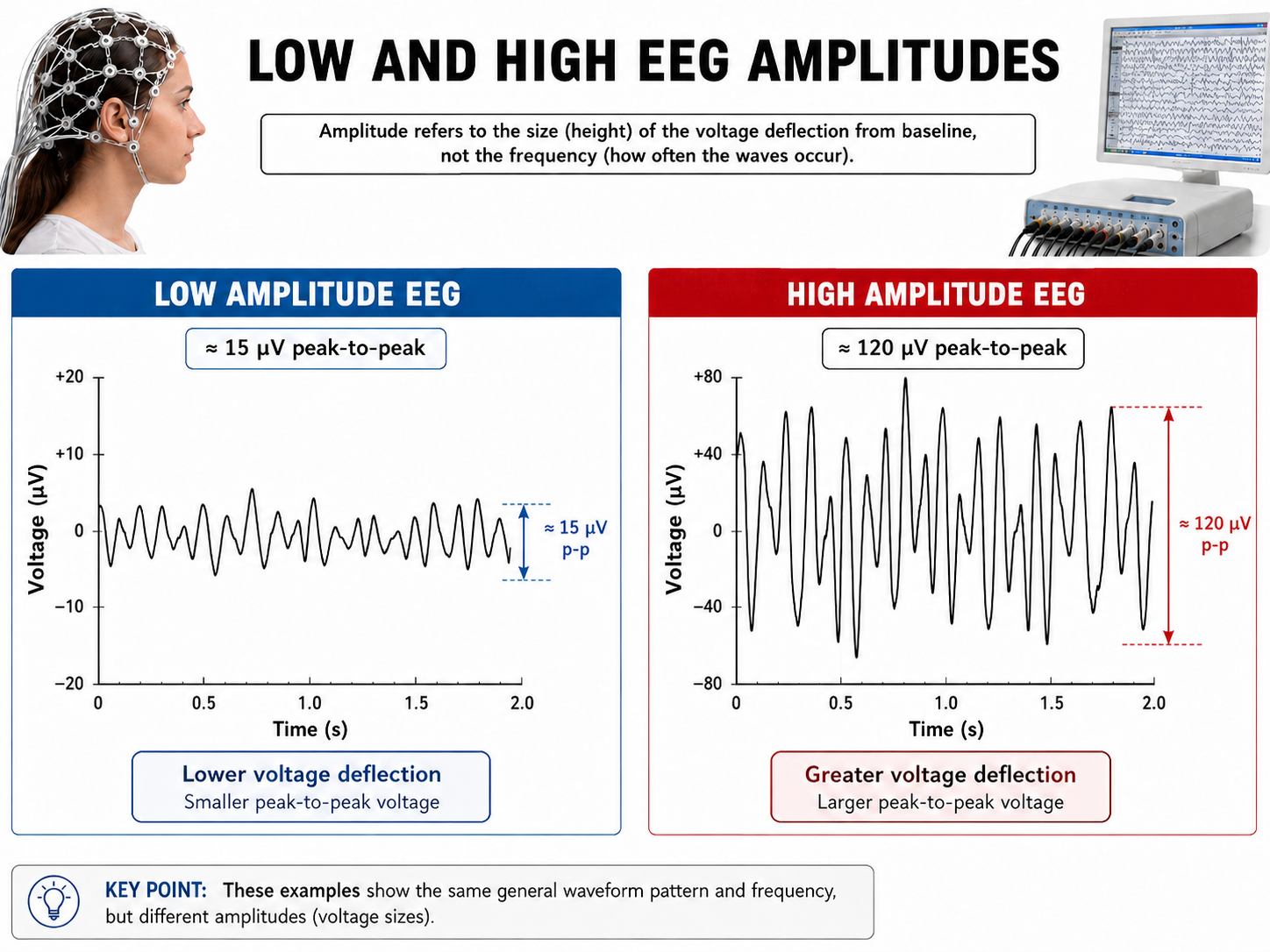
Amplitude measures the size of the voltage fluctuation and is expressed in microvolts. Amplitude is not the same quantity as power, which is proportional to the square of amplitude and is expressed in µV2.
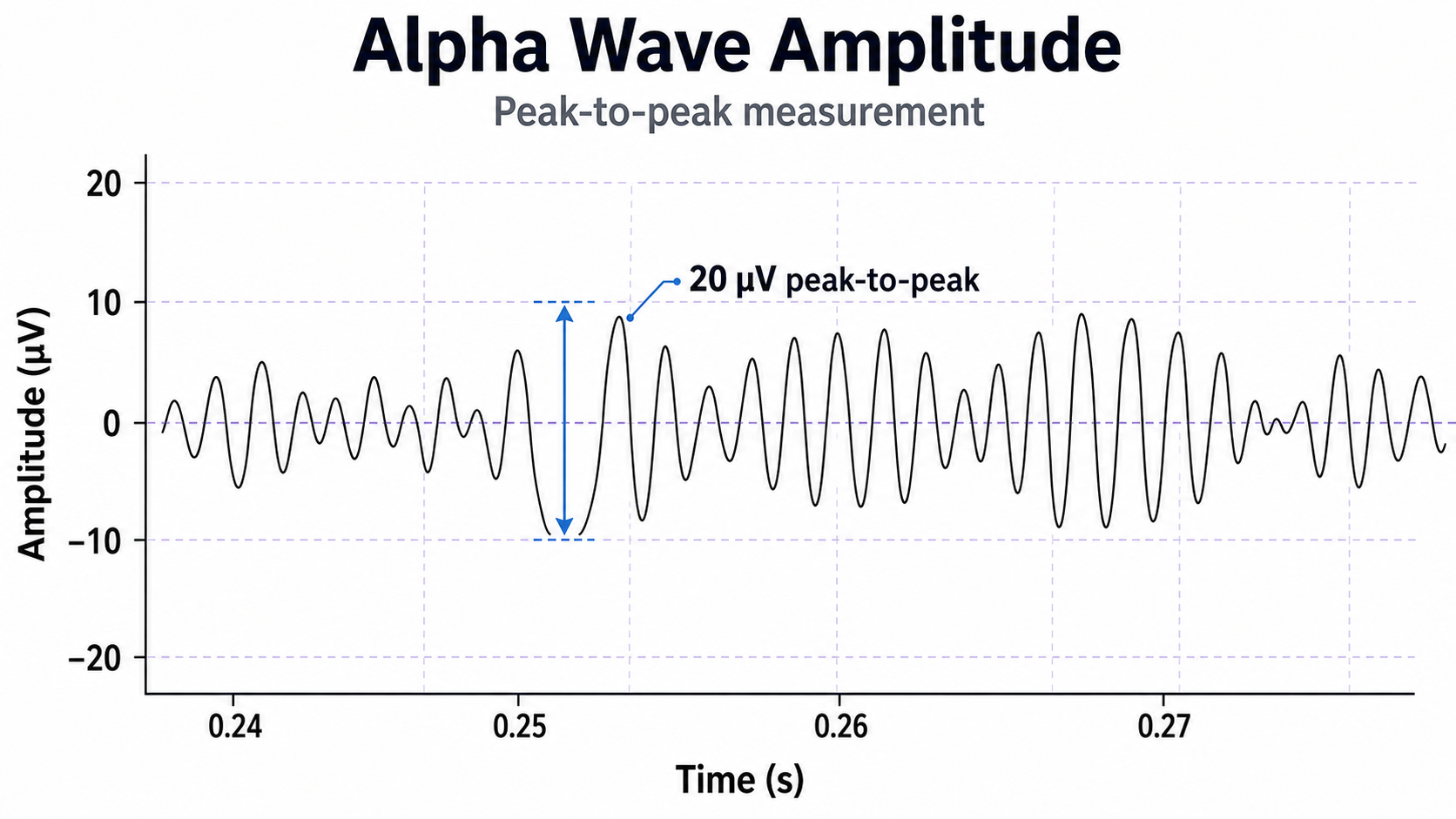
Greater synchrony in firing among neurons results in higher amplitudes detected at the scalp, as in the graphic below.
The EEG plots voltage changes over time, which can be displayed on a graph. The sampling rate is the number of measurements per second (Hz). Precision is the number of voltage gradations or steps.
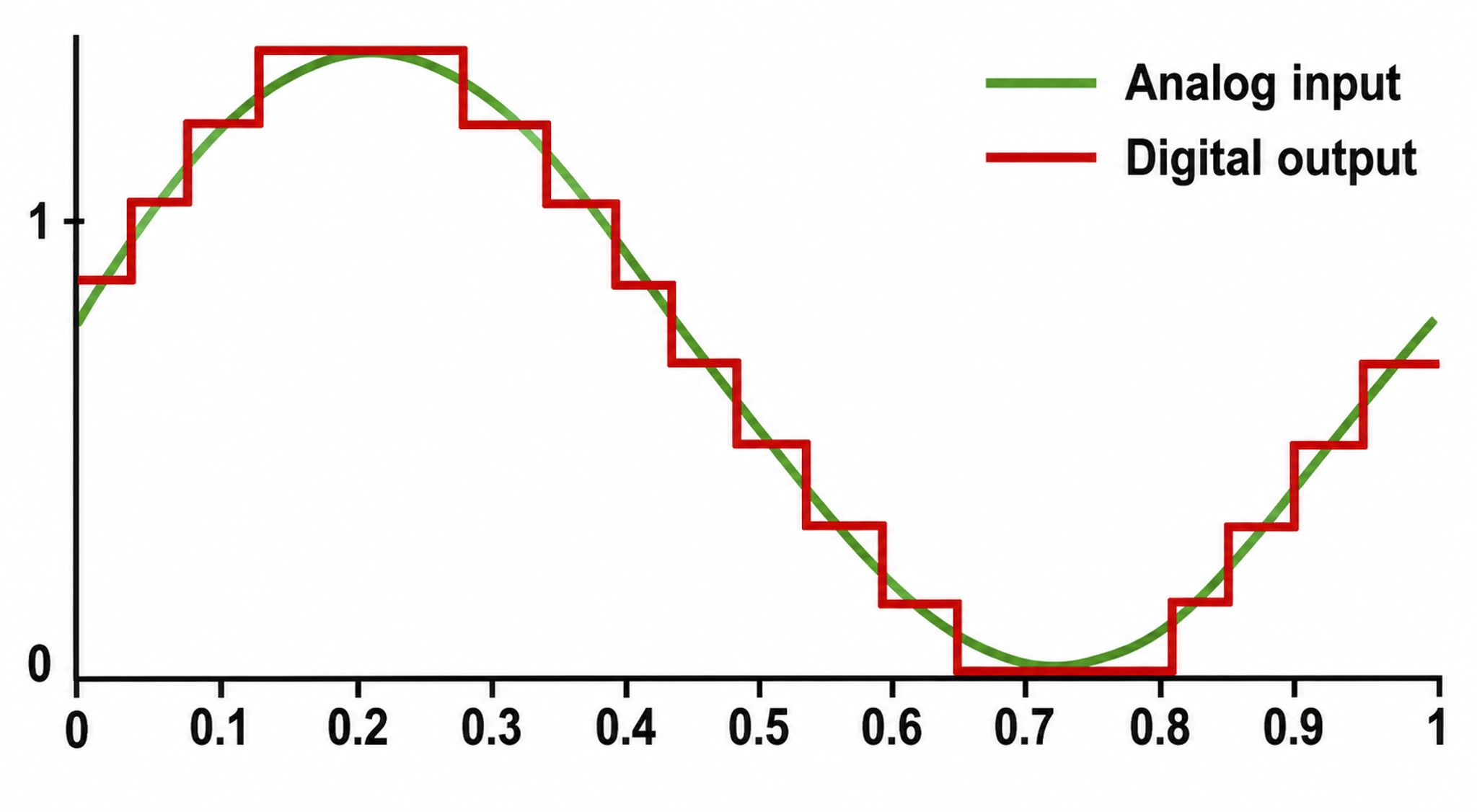
According to the Nyquist theorem, the sampling rate must be at least twice the highest frequency of interest to represent that activity accurately. Sampling too slowly causes aliasing, in which fast activity masquerades as a slower, false rhythm. Recording 30-Hz beta therefore requires sampling well above 60 Hz, and modern systems sample at 256 Hz or higher (Collura, 2014).
The analog-to-digital (A/D) converters that transform voltages into numerical values vary in precision: more bits correspond to greater accuracy. Graphic adapted from © Fouad A. Saad/Shutterstock.com.
EEG Frequencies
The raw EEG contains all EEG frequencies, just as white light contains all light frequencies. Digital filters separate the EEG frequencies just as a prism separates individual colors. Graphic © kmls/ Shutterstock.com.

EEG frequency is measured in cycles per second or Hz. Count the number of peaks or count the number of zero (0.0) crossings divided by 2.
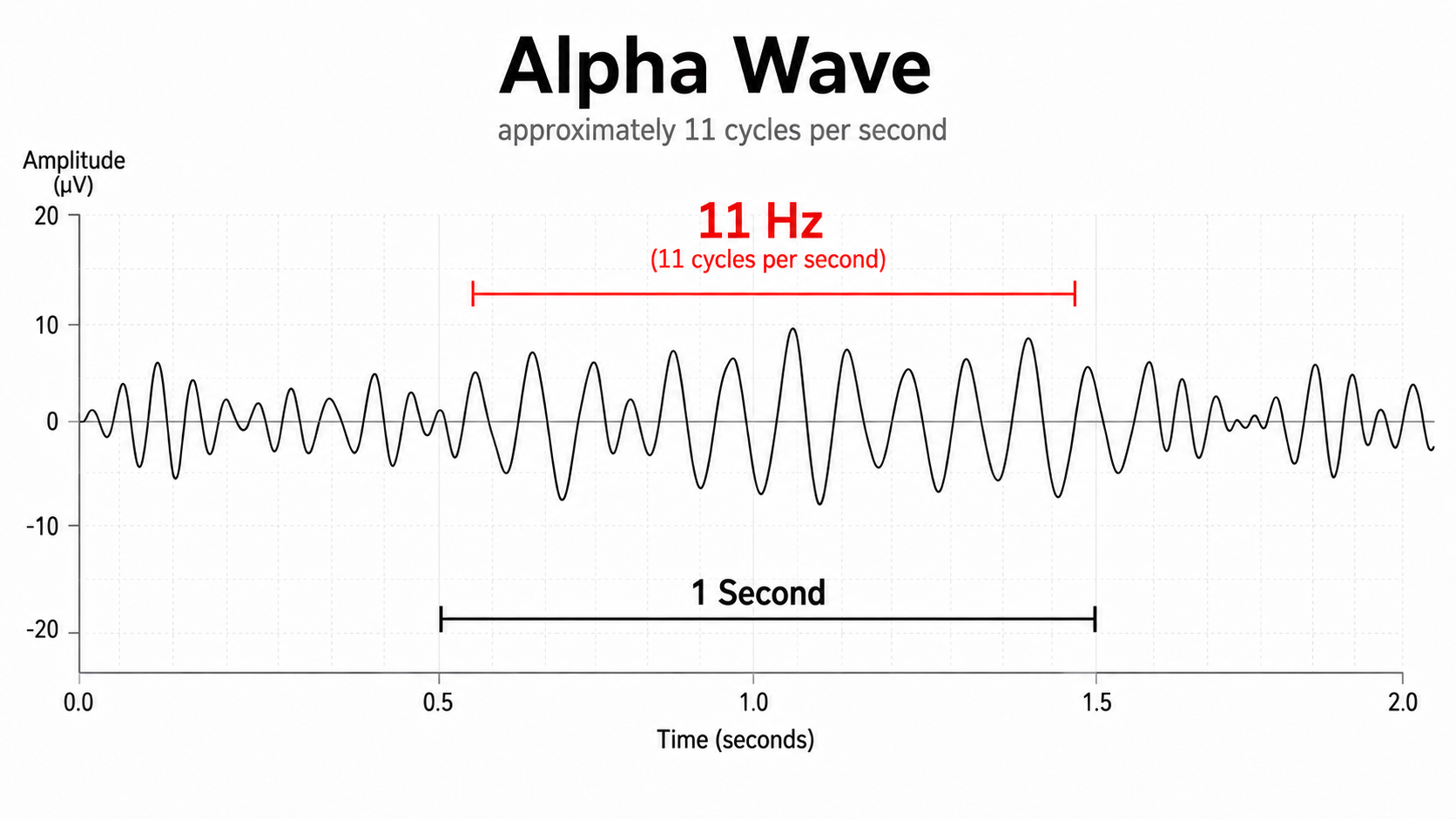
The slower the waves, the lower the EEG frequency.
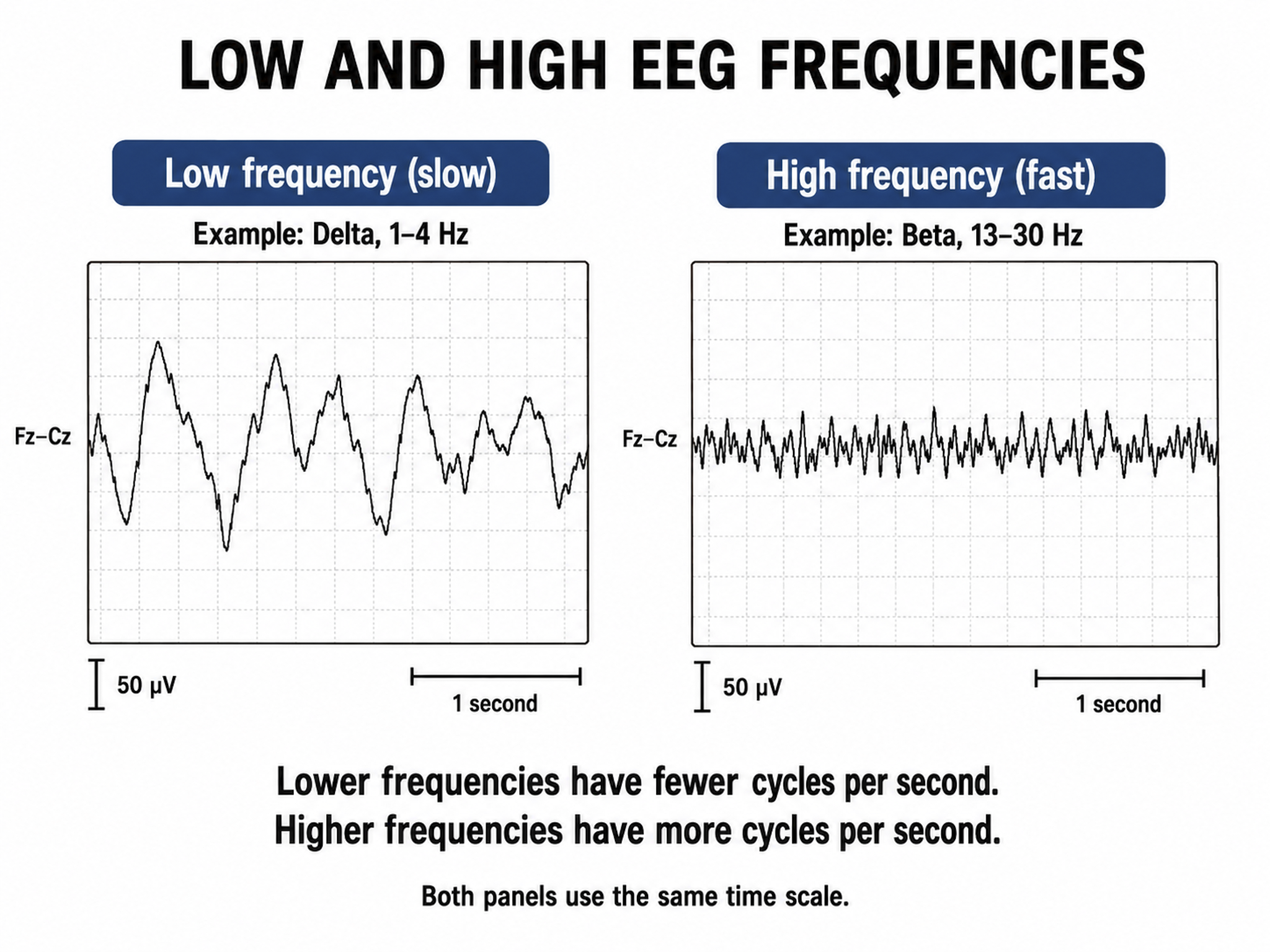
The movie is a 19-channel BioTrace+ /NeXus-32 display of EEG activity from 1-64 Hz activity broken into its component delta, theta, alpha, and beta frequency bands by digital filters © John S. Anderson.
The movie is a 19-channel BioTrace+ /NeXus-32 display of alpha activity © John S. Anderson. Brighter colors represent higher alpha amplitudes. Frequency histograms are displayed for each channel. Notice the runs of high-amplitude alpha waves.
EEG Oscillations
The generation of oscillatory activity, sometimes called spindle behavior, is likely due to the interaction between thalamocortical relay neurons (TCR), reticular nucleus neurons (RE), and interneurons. These interactions are mediated by diverse neurotransmitters, including acetylcholine and GABA.
Circuits Contributing to the EEG
Feedforward, thalamocortical, and intra-cortical networks help generate the EEG.
Spindling or Bursting Activity
Spindling is a synaptically-generated oscillation in a circuit that necessarily includes reticular nucleus neurons (RE).
The movie below is a BioTrace+ /NeXus-32 display of EEG spindling activity © John S. Anderson.
The various spindle frequencies, which have often been interpreted as reflecting different types of oscillations, merely depend on various durations of the hyperpolarizations (negative shifts) in thalamic-cortical relay neurons. Long duration hyperpolarizations, as during ... deeply EEG-synchronized states, are associated with 7 Hz or even lower-frequency spindles, while relatively short hyperpolarizations result in ... higher frequencies (14 Hz) (Steriade, 2005).
The Purpose of Oscillatory Activity
A single neuron can influence multiple postsynaptic targets located between 0.5 and 5 mm away with conduction periods of between 1 and 10 ms. This time difference becomes progressively more pronounced when more complex events involve progressively larger assemblies of neurons. It may take hundreds of thousands of neurons, stimulating multiple postsynaptic targets, for the desired outcome to occur. When this many neurons are involved, it becomes increasingly clear that there is a need for organization and structure to manage this diverse activity.
Timing is everything since action potentials arrive from a large number of sources. The nervous system must correctly register arrival times to recognize a face, recall a name, or remember personal history and context. Oscillatory activity provides the temporal framework that coordinates this precisely timed communication.
Hierarchical Processing
Complex events require that the systems involved operate within a spatial and temporal hierarchy. Each oscillatory cycle is a window of time within which processing can occur, with a beginning and an end within which encoded or transferred messages must complete their tasks. Groups of neurons, close or distant, interact most effectively when firing windows are synchronous. The brain does not operate continuously but in discontinuous packets.
Multiple Oscillators
"Oscillatory classes in the cerebral cortex show a linear progression of the frequency classes on the log scale. In each class, the frequency ranges ('bandwidth') overlap with those of the neighboring classes, so that frequency coverage is more than four orders of magnitude" (Buzsáki, 2006). Graphic adapted from Buzsáki (2006).
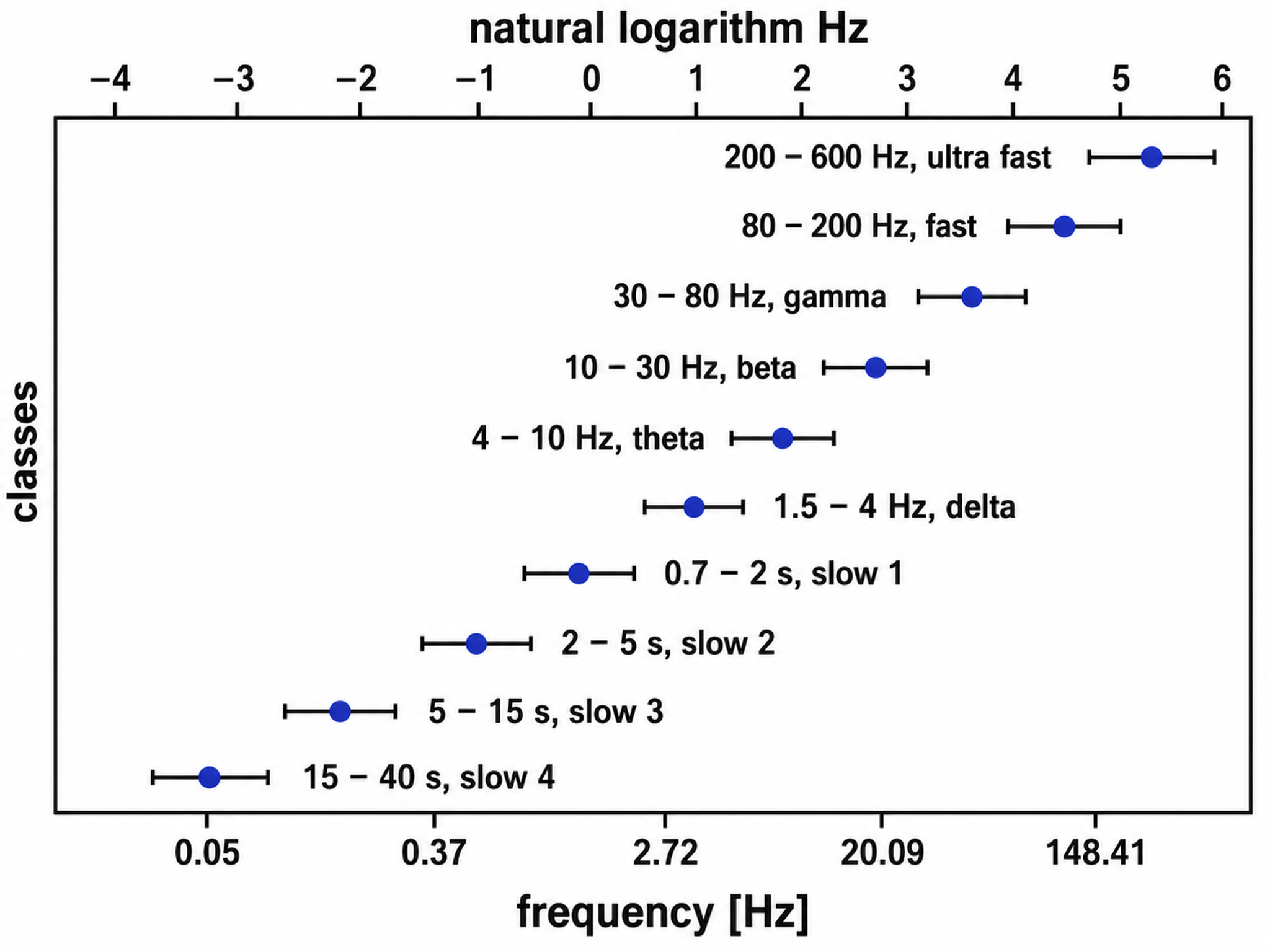
Frequency Determines Complexity
The wavelength or frequency of the EEG band determines how long the processing window will remain open and, therefore, the size of the neuronal pool involved. Because of the distances involved, longer wavelengths (slower frequencies) allow larger groups of more distant neurons to be assembled and coordinated. Different frequencies organize different types of connections and different levels of computational complexity. This principle has direct clinical relevance: training slower frequencies influences broader, more distributed brain networks, while training faster frequencies targets more localized processing.
Local Versus Global Decision-Making
Short time windows of fast oscillators facilitate local integration, primarily because of the limitations of axon conduction delays. Fast oscillations favor local decisions. Slow oscillators can involve many neurons in large and/or distant brain areas. Slow oscillations favor complex, global decisions.
Complexity Versus Frequency
The organizing variable in this literature is the cortical distance over which activity must be integrated rather than task complexity as such. Long-range fronto-parietal integration during working-memory retention appears as 4-7 Hz synchronization (Sarnthein et al., 1998). Integration between neighboring temporal and parietal cortex during multimodal object processing, whether the object arrives as a spoken word, a written word, or a picture, appears in the 13-18 Hz beta1 range (von Stein et al., 1999). Integration local to a single area appears in the gamma range (von Stein & Sarnthein, 2000).
Traveling Waves Help Coordinate Widespread Brain Networks
Zhang et al. (2018) recorded oscillations between 2 and 15 Hz whose phase gradients showed them moving across the cortex at 0.25-0.75 meters per second, and suggested that such traveling waves help coordinate brain networks at a large scale and support connectivity.
Summary of EEG Oscillations
When the CNS processes incoming content, separate areas detect features of salient content, including visual, auditory, tactile, kinesthetic, and olfactory information. The CNS shares, integrates, and compares current with previous content, analyzes it, and makes decisions regarding memory and responses. Interacting networks linked by electrical and chemical signals perform this work, and we record the electrical potentials generated by this complex and dynamic network activity as the EEG.
The movie below of bursting alpha shows the sequential synchronization/desynchronization of groups of neurons. Higher voltage bursts are followed by voltage decreasing toward zero. These voltage fluctuations reflect rhythmic changes in the local field potential. This BioTrace+ /NeXus-32 video © John S. Anderson.
Key Takeaways
The scalp EEG is generated mainly by large pyramidal neurons in cortical columns whose apical dendrites, aligned perpendicular to the surface, sum their postsynaptic potentials into local field potentials. The EEG preferentially detects radial dipoles on gyral crowns and can miss tangential sources buried in sulci, and volume conduction further blurs each source across the scalp. Amplitude reflects how many neurons fire synchronously, while frequency reflects how fast they oscillate, with slower rhythms coordinating larger and more distant networks. Generators such as the thalamus pace much of this rhythmic activity, producing familiar patterns like the posterior dominant alpha rhythm. These principles explain both the power and the spatial limitations of the signal clinicians train.
Check Your Understanding
- Why are large pyramidal neurons in cortical columns the primary generators of the scalp EEG?
- How does the orientation of a dipole, radial versus tangential, affect whether the EEG can detect it?
- What does EEG amplitude reveal about the underlying neural population, and what does frequency reveal?
- According to the Nyquist theorem, why must the sampling rate be at least twice the highest frequency of interest?
- How does volume conduction limit the spatial resolution of the scalp EEG?
Definition of ERPs and SCPs
This section defines event-related potentials (ERPs) and slow cortical potentials (SCPs), two categories of brain electrical activity with important clinical applications. ERPs reveal how the brain processes specific stimuli, while SCPs reflect broader shifts in cortical excitability that neurofeedback clinicians can train.
Sensory evoked potentials are a subset of event-related potentials (ERPs)
Event-related potentials (ERPs) represent the brain's responses to external stimuli, events, or cognitive/motor tasks. ERPs can be detected throughout the cortex, and investigators monitor them by placing electrodes at midline sites (Fz, Cz, and Pz), sometimes using dense arrays of 64 to 256 electrodes. A computer analyzes a subject's EEG responses to the same stimulus or task over many trials to subtract random EEG activity. For a fixed stimulus, task, and electrode site, the averaged ERP is highly replicable, with negative and positive peaks at characteristic latencies following the stimulus. Morphology and scalp topography differ across modalities and tasks, so there is no single ERP waveform.
Sensory evoked potentials are a subset of ERPs elicited by external sensory stimuli (auditory, olfactory, somatosensory, and visual). In the auditory modality they show a negative peak near 80-100 ms and a positive peak near 170 ms following stimulus onset; the pattern-reversal visual evoked potential instead has its dominant positive peak near 100 ms. The orienting response ("What is it?") is a sensory ERP. The N1-P2 complex in the auditory cortex reveals whether an uncommunicative person can hear a stimulus, a clinically valuable application for patients who cannot report their own perceptual experiences. N refers to a negative potential and P to a positive potential.
Motor ERPs are detected over the primary motor cortex (precentral gyrus) during movement, and their amplitude is proportional to the force and rate of skeletal muscle contraction (Thompson & Thompson, 2015).
Slow cortical potentials modulate the excitability of associated neurons
Slow cortical potentials (SCPs) are gradual changes in the membrane potentials of cortical dendrites that last from 300 ms to several seconds. SCPs are characterized by low-frequency oscillations typically below 1 Hz, distinct from other brain rhythms such as delta (1-4 Hz) and spindling (7-14 Hz). SCPs have been observed at approximately 0.3 Hz, and their depolarizing-hyperpolarizing components have been extensively analyzed.

These potentials include the contingent negative variation (CNV), the readiness potential (Bereitschaftspotential), and movement-related potentials (MRPs). Slow cortical potentials are best understood as the low-frequency subclass of event-related potentials rather than as a category separate from them. The faster P300 and N400 components described below are event-related potentials but are not slow cortical potentials (Andreassi, 2007).
SCPs modulate the firing rate of cortical pyramidal neurons by exciting or inhibiting their apical dendrites and group the classical EEG rhythms using these synchronizing mechanisms (Steriade, 2005). For neurofeedback clinicians, SCPs represent a powerful training target because they regulate the very neural excitability that underlies faster EEG rhythms.
The movie is a 19-channel BioTrace+ /NeXus-32 display of SCPs © John S. Anderson. Brighter colors represent higher SCP amplitudes. As noted earlier, which direction a negative shift travels on screen depends on the software settings; the convention in electroencephalography is to plot negative up. Negative SCPs are produced by the depolarization of apical dendrites and increase the probability of neuron firing. Positive SCPs are produced by the hyperpolarization of these dendrites and decrease the likelihood of neuron firing.
The contingent negative variation (CNV) is a steady, negative shift in potential (15 µV in young adults) detected at the vertex. This slow cortical potential may reflect expectancy, motivation, intention to act, or attention. The CNV appears 200-400 ms after a warning signal (S1), peaks within 400-900 ms in the short warning-interval paradigm shown below, and sharply declines after a second stimulus that requires a response (S2). With longer S1-S2 intervals the CNV becomes biphasic and its late component peaks just before S2.
The readiness potential is a slow-rising, negative potential (10-15 µV) detected at the vertex before voluntary and spontaneous movement. This SCP precedes voluntary movement by 0.5 to 1 second in the late phase illustrated below and peaks at about the time the subject responds. Recorded over a longer epoch, an earlier component begins roughly 1.5 to 2 seconds before movement onset (Shibasaki & Hallett, 2006). It is separate from the CNV.
Movement-related potentials (MRPs) occur at 1 second as subjects prepare for unilateral voluntary movements. MRPs are distributed bilaterally with maximum amplitude at Cz. The supplementary motor area and primary motor and somatosensory cortices generate these potentials (Babiloni et al., 2002).
P300 and N400 ERPs are classified as long-latency potentials due to their extended latencies following stimulus onset.
The P300 potential is an ERP whose peak typically falls between 250 and 500 ms after stimulus onset, lengthening with task difficulty, age, and cognitive impairment. The largest amplitude positive peaks are located over the parietal lobe. Researchers elicit the P300 by exposing subjects to an odd-ball stimulus, a meaningful stimulus that differs from others in a series (a colored playing card presented in a series of monochrome cards). The P300 may reflect an event's subjective probability, meaning, and information transmission. Research shows this is separate from the CNV (Stern, Ray, & Quigley, 2001).
Shorter P300 latencies may reflect better allocation of attention, and researchers have measured longer P300 latencies in ADD than non-ADD samples. Experimental subjects show longer latencies when lying than when telling the truth (Farwell & Donchin, 1991; Thompson & Thompson, 2015).
The N400 potential is an ERP elicited when we encounter semantic violations like ending a sentence with an incongruent word ("The handsome prince married the beautiful fish"), or when the second word of a pair is unrelated to the first (BATTLE/GIRL). Warren and McIlvane (1998) speculate that the N400 is evoked whenever a conceptual system encounters a mismatch that violates equivalence relations. Halgren and colleagues (2002) consider it an index of the difficulty of semantic processing.
A Deep Dive Into SCPs
This section traces the history of SCP research from Richard Caton's 1875 observations through modern neurofeedback applications. It examines SCP generators, the paradox of scalp negativity during neural activation, and the clinical significance of SCPs across multiple disorders.
In 1875, Richard Caton identified what may have been the first evidence of SCPs in an article in the British Medical Journal titled "The Electric Currents of the Brain."
He reported that "feeble currents of varying direction pass through the multiplier when the electrodes are placed on two points of the external surface," and that "when any part of the grey matter is in a state of functional activity, its electric current usually exhibits negative variation." Working with a mirror galvanometer, Caton recorded deflections rather than calibrated voltages, so his one-page report contains no voltage figures. Some later researchers suggested that this signaled the discovery of the "steady potential" or the DC potential of the brain, though others have noted the possibility of equipment-based artifacts in his recordings (Niedermeyer, 1999).
From the late 1800s through the early 1900s, research into brain electrical activity turned toward observations of electrical stimulation and spontaneous electrical activity in animal studies. As technology improved, the ability of researchers to identify EEG rhythms also improved. Hans Berger is famous for his description of alpha-blocking with cognitive activity, made possible partly because of his use of more sensitive equipment (Niedermeyer, 1999).
Slow Cortical Potential Generators
Several neural mechanisms and structures within the brain generate SCPs. The generation of SCPs is primarily cortical, as evidenced by their persistence even after extensive thalamic destruction and corpus callosum transection (Steriade, Nuñez, & Amzica, 1993).
SCPs have been identified in cortical neurons, the thalamus, and glial cells. Cortical neurons in layers II to VI generate slow oscillations when the thalamus is removed or when cortical tissue is studied in vitro (in an artificial environment) or in vivo (within a living organism). Thalamic reticular neurons exhibit similar slow spontaneous oscillations in vitro, and synchronized intracortical oscillations may depend on a corticothalamic network.
The source and nature of SCPs remain in dispute. The prevailing theory holds that negative SCPs result from synchronous postsynaptic potentials in the apical dendrites of cortical pyramidal cells. Others hold that SCPs are produced by glial cells within the cortex. It appears that pyramidal neurons may be the source while the glial system is the "sink" (Strehl, 2005, personal communication).
Increased neuronal activity is associated with increased outflow of potassium ions leading to increased extracellular potassium concentrations. Glial cells depolarize when extracellular potassium concentrations increase, resulting in current flows similar to typical neuronal synaptic transmissions (Speckmann & Elger, 1999). Since glial cells are widely interconnected and have extensive processes, the glial system likely contributes to the potential changes producing SCP values recorded from the scalp.
Despite ongoing debate about SCP sources, scalp SCPs clearly represent cortical excitability. SCP negativity is associated with increased cortical excitability; high cortical negativity correlates with greater likelihood of seizures (Speckmann & Elger, 1984) and migraines (Siniatchkin et al., 2000) in susceptible individuals.
SCP positivity is associated with increased cortical inhibition. Higher-than-expected positive SCPs have been noted in children with elevated blood lead levels (Otto & Reiter, 1984). Children diagnosed with ADHD show deficient SCP self-regulation skills compared with controls (Heinrich et al., 2004). SCPs have been used to monitor anesthesia depth during surgical procedures (Sebel et al., 1997) because they are excellent indicators of arousal level.
Cortical Neurons
SCPs are primarily generated by the synchronized activity of large populations of cortical neurons. The slow shifts in membrane potential reflect changes in the overall excitability of cortical networks (Birbaumer et al., 1990). Neuron graphic © SciePro/Shutterstock.com.
Thalamocortical Interactions
Interactions between the thalamus and cortex also play a significant role in generating SCPs. Through its relay and integrative functions, the thalamus modulates cortical excitability and contributes to the slow potential changes observed (Lopes da Silva, 1991). Thalamocortical graphic © Netter.
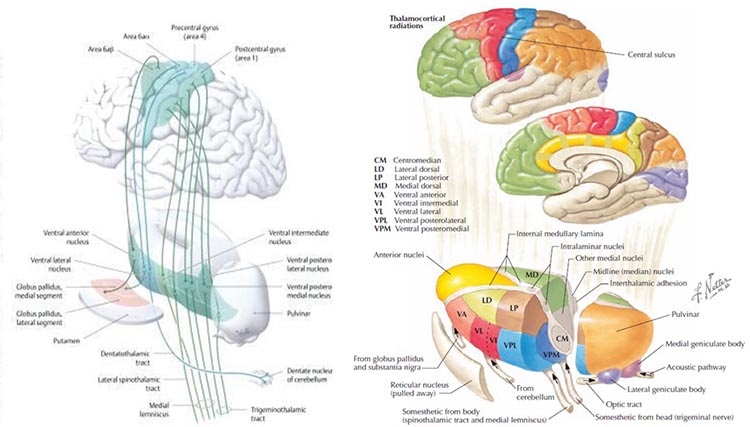
Glial Cells
Emerging evidence suggests that glial cells, particularly astrocytes, influence SCPs by modulating the extracellular environment and supporting neuronal function (Amzica & Steriade, 2002).
Glia contribute to SCPs chiefly through the potassium mechanism described above: activity-driven rises in extracellular potassium depolarize astrocytes, and because glia are extensively interconnected, that depolarization spreads as a slow field change (Speckmann & Elger, 1999). The slow oscillations of glial cells may in turn influence the timing of neuronal firing through their control of potassium ion outflow (Steriade, 2005). Astrocyte graphic © Kateryna Kon/Shutterstock.com.
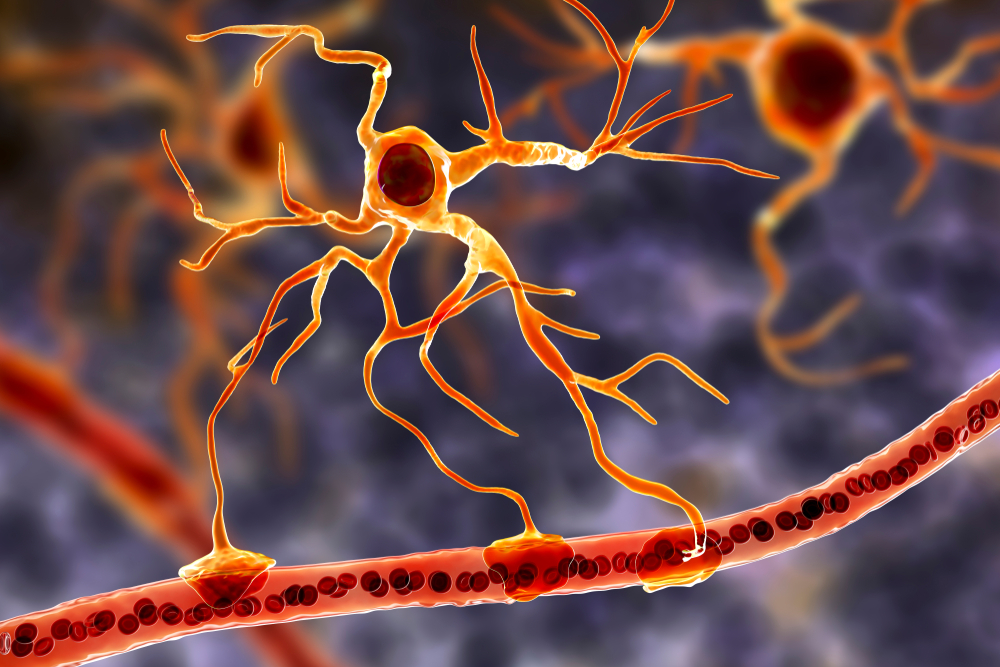
"The concept of a unified corticothalamic network that generates diverse types of brain rhythms grouped by the cortical slow oscillation is supported by EEG studies in humans" (Mölle et al., 2002).
The Meaning of SCP EEG Activity
SCPs indicate shifts in cortical excitability and are associated with various functional brain states. Surface-negative SCPs reflect synchronized depolarization of neuronal assemblies, indicating increased cortical activity. Surface-positive SCPs correspond to decreased cortical excitation, often involving inhibitory processes (Hinterberger et al., 2003).
The negative SCPs detected at the scalp during neuronal depolarization may seem counterintuitive at first.
When neurons are activated, their cell bodies become more positive internally due to the influx of positive ions. This leaves the immediate extracellular space more negative, and that negative charge is conducted through brain tissue, cerebrospinal fluid, skull, and scalp. EEG electrodes on the scalp detect this conducted negative potential, resulting in a negative deflection on the EEG trace.
This phenomenon is often referred to as paradoxical negativity in EEG literature (Birbaumer et al., 1990). What we record on the scalp is not a direct measure of neuronal membrane potential, but rather the result of complex electrical field propagation through various tissues (Elbert et al., 1980). Paradoxical positivity occurs when neurons are hyperpolarized.
This relationship is crucial for understanding the neurophysiological mechanisms underlying SCPs (Birbaumer et al., 1990). This paradoxical negativity graphic was adapted from Brienza and Mecarelli, 2019. Their original illustration is available under the license CC BY 3.0.
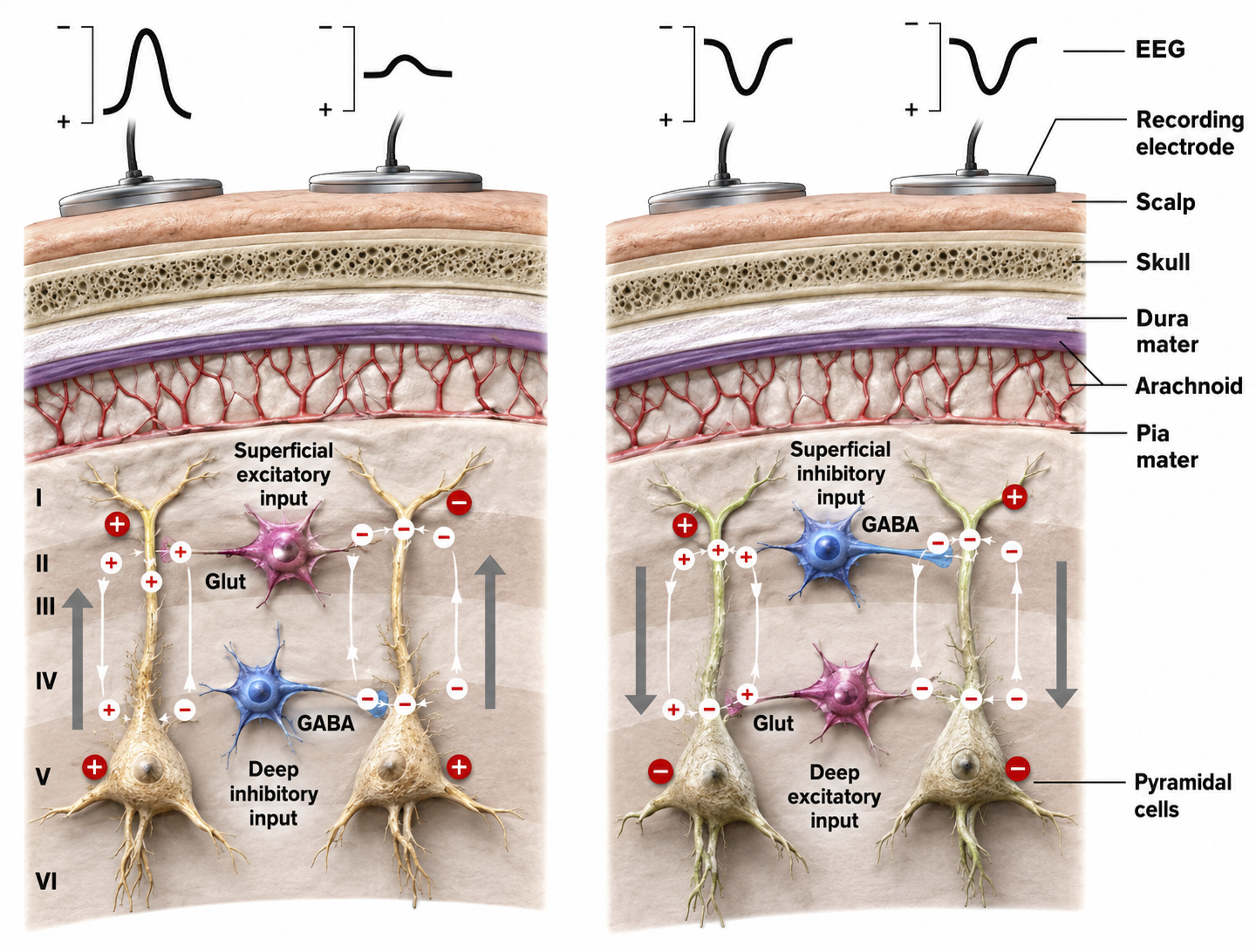
Schematic drawing of the scalp EEG registering negative (A) and positive (B) deflections elicited from summated EPSPs and IPSPs derived from pooled pyramidal cells. Cells releasing glutamate and GABA provide excitatory and inhibitory superficial and deep synaptic connections, resulting in an electrophysiological sink or source. EEG = electroencephalography; EPSPs = excitatory postsynaptic potentials; GABA = gamma-aminobutyric acid; IPSPs = inhibitory postsynaptic potentials. Figure courtesy of Anteneh Feyissa M.D. and Mayo Clinic.
Caton (1875) observed that cortical grey matter shows a negative variation in its electric current whenever it is functionally active. Underlying "tone" or valence factors determine the firing characteristics of neurons within a network. When SCPs are more positive, cortical neurons fire less due to hyperpolarization. When SCPs are more negative, firing increases due to depolarization.
SCPs participate in cognitive processes such as attention, preparation, and intention. Negative SCP shifts are linked to increased cortical excitability and readiness to respond, while positive shifts are associated with decreased excitability and relaxation (Birbaumer et al., 1990).
Some types of SCPs are event-related, including the Bereitschaftspotential (BP or readiness potential), contingent negative variation (CNV), and stimulus-preceding negativity (SPN). These represent slow negative waves related to anticipating a stimulus or preparing for a movement (Brunia et al., 2012). The BP occurs before executing a self-paced movement, CNV occurs when a preparatory stimulus foretells an imminent response demand, and SPN occurs after a movement when waiting for accuracy feedback.
The slow rhythm of SCPs is often combined with delta oscillations and these rhythms are phase-locked, suggesting close interaction between different frequency bands (Steriade, Nuñez, & Amzica, 1993).
SCPs play a crucial role in motor preparation and execution. The readiness potential (Bereitschaftspotential) precedes voluntary movements and reflects the planning and initiation of motor actions, particularly relevant for clinicians working with athletes and military personnel, where motor preparation timing can be a performance target.
SCPs are also associated with emotional and motivational states. Negative SCPs can indicate increased arousal and emotional engagement, whereas positive SCPs can reflect relaxation and disengagement (Hinterberger et al., 2003).
Psychological and Medical Disorders
SCPs have been extensively studied in various psychological and medical conditions. The following conditions illustrate the breadth of SCP-related assessment and intervention.
Attention-Deficit/Hyperactivity Disorder (ADHD)
Individuals with ADHD often exhibit abnormal SCP patterns, with a reduced ability to generate negative SCP shifts. Neurofeedback training targeting SCPs has shown promise in improving attention and reducing hyperactivity (Heinrich et al., 2004). This is one of the most well-studied applications of SCP neurofeedback.
Epilepsy
SCP neurofeedback has been explored as a treatment for epilepsy. Training individuals to increase positive SCP shifts can reduce cortical excitability and decrease seizure frequency (Rockstroh et al., 1993).
Parkinson's Disease
Studies have shown that patients with Parkinson's disease (PD) exhibit abnormal SCP patterns, particularly during motor tasks (Brittain & Brown, 2014). These abnormalities include altered amplitude and timing of SCPs, associated with impaired initiation and execution of voluntary movements.
During NREM sleep, cortico-basal slow wave delta activity increases while beta activity decreases. Deep brain stimulation (DBS) further modulates this altered activity, enhancing cortical delta and reducing alpha and low beta power. These findings suggest that SCPs and their interaction with other brain rhythms are significantly altered in PD, contributing to sleep dysfunction and spontaneous awakenings (Anjum et al., 2023).
SCPs are used to monitor the effects of therapeutic interventions such as DBS on cortical function in PD patients.
SCP neurofeedback has shown potential as a complementary treatment for PD, aiming to train self-regulation of brain activity associated with motor control.
Research suggests that SCP neurofeedback can improve motor function in PD patients (Kober & Wood, 2014). Some studies report better control over tremors and rigidity, along with improvements in non-motor symptoms including mood and cognitive function.
While promising, the current evidence is based on limited studies with small sample sizes. More extensive clinical trials are needed to establish long-term efficacy and generalizability.
Depression
SCP abnormalities are observed in depression, with patients often showing reduced amplitude of SCP shifts. Neurofeedback interventions aiming to normalize SCP patterns have shown potential in alleviating depressive symptoms (Strehl et al., 2017).
Sleep
SCPs play a role in sleep regulation and quality, making them relevant to clinicians who work with sleep-related complaints.
Sleep Onset and Maintenance
SCPs are involved in the transition from wakefulness to sleep. Positive SCP shifts are associated with sleep initiation and maintaining sleep stability (Sterman, 1996).
Sleep Disorders
SCPs are closely linked to sleep rhythms, particularly during NREM sleep (Anjum et al., 2023). The slow oscillations of SCPs facilitate the synchronization of neuronal activity essential for restorative sleep functions. In PD, the suppression of slow waves and the increase in subcortical beta activity before spontaneous awakenings highlight the critical role of SCPs in maintaining sleep quality.
Abnormal SCP patterns have been linked to insomnia. Neurofeedback training targeting SCPs can improve sleep onset latency and enhance overall sleep quality (Hoedlmoser et al., 2008).
Performance
Enhancing SCP activity through neurofeedback training has improved performance in various cognitive and motor tasks. This makes SCP training relevant not only for clinical populations but also for optimal performance programs serving athletes and military personnel.
Cognitive Performance
SCP training can enhance attention, memory, and executive function, likely due to improved cortical excitability regulation and better cognitive state management (Vernon et al., 2003).
Motor Performance
SCP neurofeedback improves motor performance, particularly in tasks requiring precise timing and coordination, attributed to the role of SCPs in motor preparation and execution (Gruzelier et al., 2014).
SCP Research
Research into human brain electrical characteristics became primarily focused on phasic phenomena from AC-coupled recordings, a trend continuing today with neurofeedback focusing primarily on training AC frequencies in the 1 to 60 Hz range.
The study of SCPs continued in physiology and animal research. Only recently has interest increased in observing SCP values in the human EEG and correlating them with cognitive activity, sensory processing, and motor activity. SCPs are distinguished from short-latency ERPs up to 500 ms and reflect cortical processes requiring more than one second to complete. Such changes occur in task-specific cortical areas and can be displayed using topographic maps, with areas of activation showing surface negative potential changes (Altenmuller & Gerloff, 1999).
Operant conditioning of SCP changes is an even more recent study area. One reason for increased interest is the excellent work by Birbaumer and colleagues (1999) at the University of Tübingen, demonstrating that SCPs can be operantly conditioned with positive outcomes for a variety of disorders. The recent availability of DC-coupled amplifiers has also contributed to this interest (Altenmuller & Gerloff, 1999).
According to Niedermeyer (1999), "DC" can mean several things. DC means direct current, a current without oscillations. From an electrophysiological perspective, "DC shifts" are ultra-slow potentials below the typical EEG oscillation frequency, generally around 0.1-0.2 cycles per second but potentially extending up to 1 cycle per second. So SCPs are not true direct current, though their oscillations are so slow that they are "DC-like" phenomena.
DC also refers to "direct coupling" (Niedermeyer, 1999) and describes an amplifier type that does not use capacitors between amplification stages and uses an infinite time constant for optimal DC recording. Until recently, this was difficult to achieve for EEG recording. Most conventional EEG amplifiers use capacitors in the input stage, which reject DC voltages and create a finite time constant that interferes with access to DC phenomena.
An approximation of DC information can be obtained from an AC amplifier by using a rectifier or extending the time constant to approximately 10 seconds (Kotchoubey et al., 1999). A thorough discussion of amplifier characteristics is beyond the scope of this article. Several excellent chapters on this subject can be found in Niedermeyer and Lopes da Silva (1999).
Recent studies have used SCPs to evaluate various task-oriented responses. Birbaumer and colleagues trained SCPs to reduce seizures (Daum et al., 1993; Kotchoubey et al., 1997, 1999, 2001, 2002), and other groups applied SCP feedback training to improve ADHD (Heinrich et al., 2004; Strehl, 2004, personal communication) and schizophrenia (Schneider et al., 1992).
SCP feedback training targets general arousal characteristics using a single measure, compared to other EEG training approaches that reward increases and/or decreases in certain frequency combinations. SCP feedback may provide a less complex approach to training neuronal activity, potentially offering greater accessibility through clinician-supervised home training devices.
Most research to date has used the Cz electrode site. However, at least one investigation trained left hemisphere language sites, demonstrating improved word processing following negativity training and diminished performance following positivity training (Pulvermüller, 2000). Studying the effects of SCP training at other electrode sites would be a productive research direction.
Some efforts have identified SCP values using multiple electrodes in a quantitative EEG paradigm. Basile and colleagues (2004) used four 32-channel DC-coupled amplifiers to compare SCP responses in schizophrenic patients with normal controls. They found significant variations, with controls showing simply-organized positivity and negativity patterns, while schizophrenic patients showed much more complex, fragmented patterns of activation and inhibition.
There are only a few clinically available DC-coupled amplifiers capable of accurately monitoring SCP activity. An Internet search yielded several research-grade devices with high prices and a couple of devices priced for clinical practice. A new 32-channel DC-coupled device for quantitative EEG assessments has also recently been released.
One attraction of DC amplifiers is the capability to monitor and/or train both SCPs and typical EEG frequencies. DC amplifiers are optimized for SCP and can also record faster frequencies, particularly those with better analog-to-digital conversion characteristics (bit size, not sampling rate).
Higher A/D conversion values (more bits per sample) allow newer DC amplifiers to resolve microvolt-level slow cortical potentials while simultaneously accommodating electrode offset potentials of tens of millivolts without saturating. The offset is what occupies the millivolt range; the slow cortical potentials themselves remain microvolt signals.
The training of SCP shifts is a fairly new endeavor. Much remains to be learned about the effects of training both positivity and negativity conditions at various electrode sites for individuals with diverse presenting concerns and neurophysiological characteristics.
This author's recent clinical experiences suggest that training SCP using newer, more accurate amplifiers may result in more pronounced changes occurring more quickly. Thus, it will be important to develop protocols with more specificity and flexibility to meet the needs of diverse client populations while considering changes in equipment and software characteristics that may affect skill acquisition rates and outcomes.
Training SCP
This section reviews approaches to training slow cortical potential shifts, from early event-related paradigms to modern infra-low frequency methods.
Various approaches to training slow cortical shifts have been applied. Early research (Birbaumer et al., 1990; Birbaumer, 1999) showed a correlation between cortical negativity and reaction time, signal detection, and short-term memory. This was identified through evoked and event-related potential research, leading to training paradigms involving sequences of 8-second trials that trained both positive and negative cortical gradient shifts using real-time visual and/or auditory feedback.
When greater cortical positivity was the goal, more positive shift trials were provided, and vice versa. Additional transfer trials without feedback tested skill acquisition. This approach was the primary paradigm during the early years of SCP research (Strehl, 2009).
Other clinicians and researchers, including Susan and Siegfried Othmer and Mark Smith, addressed slow gradient shifts with training called Infra-Low Frequency Neurofeedback (Othmer, 2020) and Infraslow Neurofeedback (Smith, 2013).
Post-traumatic stress, anxiety, and other conditions involving excessive cortical activation have been addressed by training to increase overall cortical positivity, using a 4-channel approach that rewards gradual shifts in the cortical gradient through proportional audio feedback. This has resulted in several client self-reports of an altered state characterized by decreased cognitive activity while retaining awareness; the positive shift in cortical gradient appears to correspond with reduced conscious thought while preserving self-awareness (multiple clinical observations shared with John Anderson).
Conclusion
Slow cortical potentials are characterized by low-frequency oscillations typically below 1 Hz, with significant depolarizing-hyperpolarizing components. Occurring at approximately 0.3 Hz, these potentials are crucial indicators of cortical excitability and are associated with various cognitive and motor processes. Generated by cortical neurons, thalamocortical interactions, and glial cells, SCPs reflect shifts in cortical excitability linked to attention, motor preparation, and emotional states. Negative SCP shifts indicate increased excitability and readiness to respond, while positive shifts are associated with relaxation and decreased excitability.
SCPs play roles in psychological and medical disorders like ADHD, epilepsy, and depression, and are vital in sleep regulation and performance enhancement. SCP neurofeedback shows promise in improving symptoms and cognitive functions across these conditions, making it an increasingly important tool in the neurofeedback clinician's repertoire.
The author would like to thank Ute Strehl of the University of Tübingen in Germany, Dave Siever of Mind Alive Inc. in Canada, and Erwin Hartsuiker of Mind Media BV in the Netherlands for technical assistance in preparing this article.
Clinical Application
Consider a child referred for inattention and impulsivity. Children with ADHD often show a reduced ability to generate the negative slow cortical potential shifts that support cortical excitability and readiness to respond (Heinrich et al., 2004). Using SCP neurofeedback, the clinician rewards the child for producing and controlling these gradient shifts across repeated trials, including transfer trials without feedback that test whether the skill generalizes. Because SCP training targets a single, global measure of arousal regulation rather than a specific frequency band, it can offer an accessible entry point for young clients. Gains in attention and self-regulation reported in controlled trials illustrate how a slow, subthreshold cortical signal can become a practical training target.
Key Takeaways
Event-related potentials are averaged, time-locked responses to specific stimuli or tasks, whereas slow cortical potentials are gradual shifts in dendritic membrane potential lasting from hundreds of milliseconds to seconds. Negative SCP shifts reflect increased cortical excitability and readiness to respond, while positive shifts reflect reduced excitability and relaxation. Although the scalp records negativity during neuronal activation, this apparent paradox follows from how the extracellular field is conducted to the surface. SCP abnormalities appear across ADHD, epilepsy, depression, Parkinson's disease, and sleep disorders, and SCPs can be operantly conditioned. This makes SCP self-regulation a flexible target for both clinical treatment and performance enhancement.
Check Your Understanding
- How do event-related potentials differ from slow cortical potentials in their timing and in how they are measured?
- What do negative and positive SCP shifts indicate about cortical excitability?
- Why does the scalp often register negativity when cortical neurons are activated?
- Name three clinical conditions in which SCP self-regulation has been studied, and describe the general training goal in each.
- Why is the ability to operantly condition SCPs important for neurofeedback?
Neuroplasticity (LTD and LTP)
Neuroplasticity is the brain's ability to remodel its structure and function in response to experience, and it is the foundational mechanism that makes all neurofeedback training possible. Without it, the operant conditioning of brainwave activity would have no lasting effect. This section describes the levels at which neuroplastic change occurs and the two synaptic processes most central to learning, long-term depression (LTD) and long-term potentiation (LTP).
Neuroplasticity is responsible for learning and memory. Memory storage remodels neurons through changes in synaptic transmission, interneuron modulation, the formation of new synapses, and the rewiring of neural pathways (Bear et al., 2020). Because neurofeedback rewards moment-to-moment changes in central nervous system activity, it is itself a form of operant conditioning, and animal studies show that operant conditioning can induce astrogliogenesis (the creation of new astrocytes) and neurogenesis (the creation of new neurons) in structures such as the medial prefrontal cortex and hippocampus (Kerson et al., 2025; Rapanelli et al., 2011).
The graphic by Rebeca Cuesta is licensed under the Creative Commons Attribution-Share Alike 4.0 International license.
Levels of Neuroplastic Change
Neuroplastic change is not a single event but a cascade that unfolds across four nested levels: molecular, genetic, structural, and functional. The deeper levels make the more visible ones possible, so a durable training outcome reflects work at all four. Keeping these levels in mind helps clinicians explain why neurofeedback effects build gradually and why consistent practice matters.
At the molecular level, a rewarded shift in brain activity strengthens active synapses by trafficking receptors, activating calcium-dependent enzymes, and releasing growth factors such as brain-derived neurotrophic factor (BDNF). This strengthening is balanced locally, so that when some synapses are reinforced, neighboring synapses weaken to prevent runaway excitation, a process that depends on the regulatory protein Arc (El-Boustani et al., 2018). These molecular events are the first physical trace of what a client learns during a session.
At the genetic level, neural activity switches on immediate early genes within minutes, and their protein products convert short-lived synaptic changes into stable ones (Bear et al., 2020). This activity-dependent gene expression is why a single session produces only transient effects, while repeated sessions consolidate lasting change. It is the biological reason neurofeedback is delivered as a course of training rather than a one-time procedure.
At the structural level, neurons grow, prune, and reshape dendritic spines, add new connections through synaptogenesis, and extend dendritic branches (Breedlove & Watson, 2023). In some regions, the operant conditioning that drives training also supports neurogenesis and the formation of new glial cells (Rapanelli et al., 2011). These remodeled connections are the lasting hardware behind a stabilized training gain.
At the functional level, these changes alter how circuits behave, shifting synaptic efficacy, the balance of excitation and inhibition, and the connectivity between regions (Collura, 2014; Thompson & Thompson, 2015). Self-regulation of cortical rhythms can produce measurable, durable changes in cortical excitability; in one study, 30 minutes of alpha-suppression neurofeedback raised corticospinal excitability and reduced intracortical inhibition for at least 20 minutes afterward (Ros et al., 2010). Reviews of the field describe neurofeedback as a closed-loop method that reshapes the targeted networks through this kind of self-directed plasticity (Sitaram et al., 2017).
Whether training a veteran with PTSD to reduce excessive high-beta activity or an athlete to optimize alpha and theta activity, the underlying mechanism is neuroplastic change working from the molecular level upward. To learn more about neuroplasticity, view the Khan Academy video Neuroplasticity.
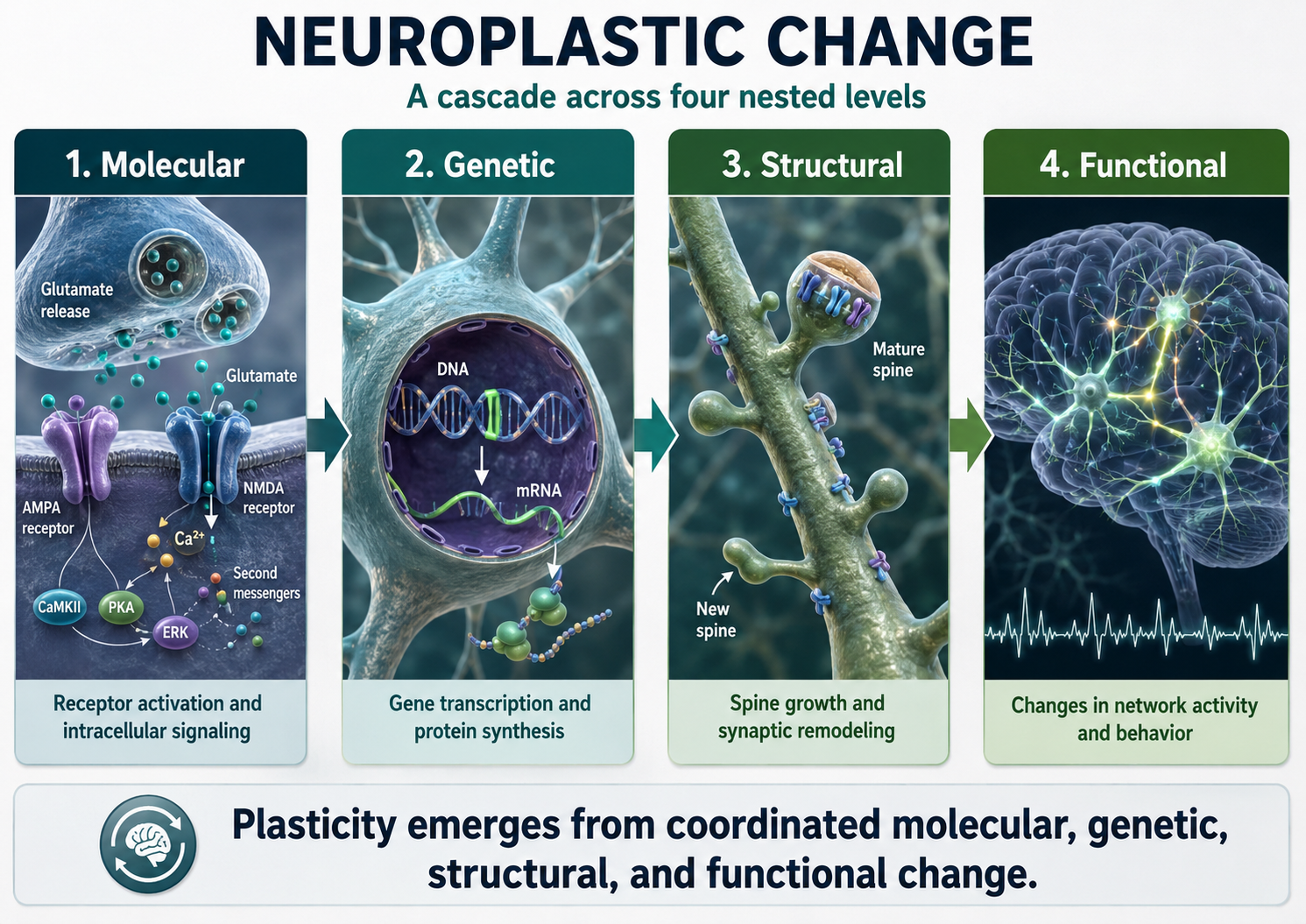
Long-Term Depression and Long-Term Potentiation
Two synaptic processes carry out functional plasticity: long-term depression and long-term potentiation. Together they provide the bidirectional control that learning requires, allowing the brain to both weaken and strengthen specific connections.
In long-term depression (LTD), synaptic transmission that coincides with slight depolarization of the postsynaptic neuron weakens the synapse, and low-frequency stimulation of afferent neurons reduces their response to future input. LTD is not simply forgetting. It is an active process that prunes less-used connections to keep neural networks efficient.
In long-term potentiation (LTP), synaptic transmission that coincides with strong depolarization strengthens the synapse, producing a stable increase in effectiveness that can persist for weeks or longer. LTP creates new synapses, enhances existing ones, and builds new dendritic branches and spines (Breedlove & Watson, 2023). Together, LTD and LTP are the cellular mechanisms through which neurofeedback training produces lasting changes in brain function.
To learn more, watch the Khan Academy video Long Term Potentiation and Synaptic Plasticity.
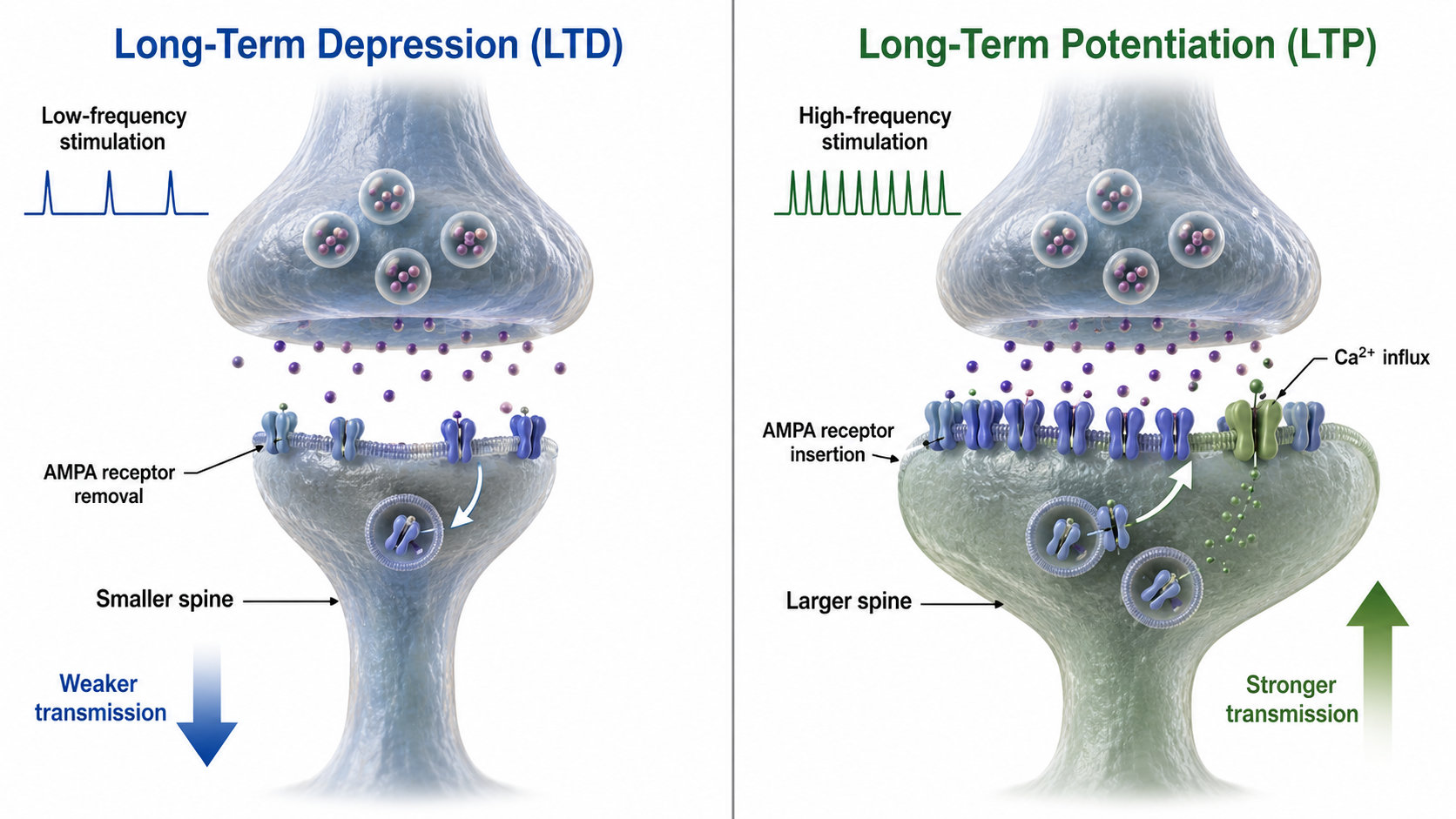
Integration: Why Neuroplasticity Matters for Neurofeedback
Neuroplasticity is the thread that connects every topic in this chapter, from the postsynaptic potentials that generate the EEG to the slow cortical shifts that clients learn to control. The four levels of neuroplastic change describe one continuous process in which a rewarded moment of brain activity leaves a molecular trace, recruits the gene expression that stabilizes it, reshapes synapses and dendrites, and finally alters how whole circuits behave. Long-term depression and long-term potentiation are the synaptic engines of that process, giving the brain the bidirectional control it needs to prune unhelpful connections and reinforce useful ones. Together these mechanisms explain how a stream of feedback tones and visual rewards becomes a lasting change in cortical function.
For the clinician, this account reframes neurofeedback as the deliberate engineering of neuroplastic change through operant conditioning (Kerson et al., 2025; Sitaram et al., 2017). Because the deeper molecular and genetic levels make the visible functional gains possible, durable outcomes depend on consistent practice across many sessions rather than a single dramatic result. The same principle explains why training effects build gradually, why consolidation continues between sessions, and why generalization to daily life reflects genuine structural remodeling rather than a temporary state (Ros et al., 2010). Whether the goal is reducing excessive high-beta activity in a veteran with PTSD or optimizing alpha and theta activity in a performing athlete, the clinician is guiding the brain through plasticity from the molecular level upward.
Key Takeaways
Neuroplasticity is the brain's capacity to remodel its structure and function with experience, and it is the mechanism that makes the effects of neurofeedback last. Change unfolds across four nested levels, molecular, genetic, structural, and functional, so that a rewarded moment of activity leaves a trace that gene expression stabilizes, synapses and dendrites embody, and circuits express as altered behavior. Long-term depression and long-term potentiation supply the bidirectional control that lets the brain weaken unhelpful connections and strengthen useful ones. Because neurofeedback is a form of operant conditioning that drives this cascade, durable outcomes depend on consistent practice that allows molecular and genetic changes to consolidate into stable structural and functional gains.
Check Your Understanding
- How do the four levels of neuroplastic change, molecular, genetic, structural, and functional, build on one another to produce a durable neurofeedback outcome?
- Why does activity-dependent gene expression help explain why neurofeedback is delivered as a course of training rather than a single session?
- How do long-term depression and long-term potentiation together provide the bidirectional control that learning requires?
- In what sense is neurofeedback a form of operant conditioning, and how does that connect to the neurogenesis and astrogliogenesis reported in animal studies?
- How would you explain to a client why neurofeedback effects build gradually and why consistent practice between and across sessions matters?
Cutting-Edge Topics in Neurophysiology
Silent Synapses in the Adult Brain
One of the most striking recent findings is the discovery of silent synapses on filopodia in the adult mouse brain (Vardalaki et al., 2022). Because these dormant connections can be recruited without disturbing existing synapses, they may reconcile the brain's competing needs for stability and flexibility. Establishing whether the adult human brain contains silent synapses could reshape how we understand learning-based interventions in mature clients.
Traveling Waves Coordinate Distant Networks
Zhang and colleagues (2018) reported that oscillations between roughly 2 and 15 Hz travel across the human neocortex as waves moving at about a quarter to three-quarters of a meter per second. Rather than treating rhythms as standing oscillations confined to one region, this work frames them as propagating signals that coordinate activity across distant areas. Traveling waves offer a fresh way to think about how large-scale connectivity is organized in time, with implications for how network-level training effects spread.
Neurofeedback as Closed-Loop, Self-Directed Plasticity
Contemporary reviews describe neurofeedback as a closed-loop method in which a person uses real-time feedback to reshape targeted brain networks through self-directed plasticity (Sitaram et al., 2017). Supporting this view, a single session of alpha-suppression training raised corticospinal excitability and reduced intracortical inhibition for at least twenty minutes afterward (Ros et al., 2010). Framing neurofeedback as deliberate, closed-loop engineering of neuroplastic change helps explain why consistent practice consolidates lasting gains.
Glossary
40-Hz rhythm: gamma rhythm hypothesized to be associated with feature binding (linking an apple's color to its shape) and attributed to the neocortex and thalamocortical neurons.
absolute refractory period: the brief interval during and just after an action potential when inactivated sodium channels make it impossible to fire a second action potential regardless of stimulus strength. It enforces one-way propagation and limits the maximum firing rate.
acetylcholine: an amine neurotransmitter that binds to nicotinic and muscarinic ACh receptors.
acetylcholine esterase (AChE): the enzyme that deactivates ACh.
AChE-R: an abnormal form of acetylcholine esterase (AChE) may render dendrites with acetylcholine receptors more excitable when stressed.
action potential: a propagated electrical signal that usually starts at a neuron's axon hillock and travels to presynaptic axon terminals.
adenylate cyclase: at a metabotropic receptor, an enzyme that transforms ATP into the second messenger cyclic AMP.
afferent: a neuron that transmits sensory information towards the central nervous system or from one region to another.
all-or-none law: once an action potential is triggered in an axon, it is propagated, without decrement, to the end of the axon. The amplitude of the action potential is unrelated to the intensity of the stimulus that triggers it.
allocortex: cortex that contains three or four layers and is comprised of the olfactory system and hippocampus.
alpha blocking: arousal and specific forms of cognitive activity may reduce alpha amplitude or eliminate it entirely while increasing EEG power in the beta range.
alpha rhythm: 8-12-Hz activity that depends on the interaction between rhythmic burst firing by a subset of thalamocortical (TC) neurons that are linked by gap junctions and rhythmic inhibition by widely distributed reticular nucleus neurons. Researchers have correlated the alpha rhythm with "relaxed wakefulness." Alpha is the dominant rhythm in adults and is located posteriorly. The alpha rhythm may be divided into alpha 1 (8-10 Hz) and alpha 2 (10-12 Hz).
alpha spindles: regular bursts of alpha activity.
alpha-subunit: a subunit of a G protein associated with the neuron membrane that breaks away to activate enzymes within the neuron when a ligand binds to a metabotropic receptor.
amino acid neurotransmitters: the oldest family of transmitters. These molecules bind to ionotropic and metabotropic receptors, so they transmit information and modulate neuronal activity. In the brain, most synaptic communication is accomplished by glutamate (generally excitatory) and GABA (generally inhibitory).
AMPA (glutamate) receptors: ionotropic receptors which open sodium channels, depolarize the neuron's membrane (producing an EPSP), and dislodge a Mg+ ion that blocks an adjacent NMDA (glutamate) receptor's calcium channel. AMPA receptors are responsible for most activity at glutamatergic synapses.
amplitude: the size of the voltage fluctuation in the EEG signal, measured in microvolts. Amplitude is not power; power is proportional to the square of amplitude and is expressed in µV2.
amygdala: the limbic system structure that participates in evaluating whether stimuli are salient (rewarding or threatening), establishing unconscious emotional memories, learning conditioned emotional responses, and producing anxiety and fear responses.
anion: a negative ion, for example, chloride (Cl-).
anterior: near or toward the front of the head, for example, the anterior cingulate.
anterior cingulate: a division of the prefrontal cortex that plays a vital role in attention and is activated during working memory. It mediates emotional and physical pain, and has cognitive (dorsal anterior cingulate) and affective (ventral anterior cingulate) conflict-monitoring components.
anterior commissure: a bundle of nerve fibers that crosses the midline and connects the left and right temporal lobes and the hippocampus and amygdala.
apical dendrite: a dendrite that arises from the top of the pyramid and extends vertically to layer 1 of the neocortex.
arousal: a process that combines alertness and wakefulness, produced by at least five neurotransmitters, including acetylcholine, histamine, hypocretin, norepinephrine, and serotonin.
aspinous (smooth) neurons: neurons without dendritic spines that are believed to be inhibitory.
astrocytes: star-shaped glial cells that communicate with and support neurons and help determine whether synapses will form.
asynchronous waves: neurons depolarize and hyperpolarize independently.
ATP: energy source for a neuron's sodium-potassium transporters.
autoreceptors: metabotropic receptors that can be located on the membrane of any part of a neuron. They detect neurotransmitters the neuron releases, generate IPSPs that inhibit the neuron from reaching the excitation threshold, and regulate internal processes like transmitter synthesis and release through the second messenger system.
axoaxonic synapses: junctions between two axons that do not affect the generation of an action potential, only the amount of neurotransmitter distributed.
axodendritic synapses: junctions between axons and dendrites that determine whether the axon hillock will initiate an action potential.
axon: long, cylindrical structures that convey information from the soma to the terminal buttons. An axon also transports molecules in both directions along the outer surface of protein bundles called microtubules.
axon hillock: a swelling in the cell body where a neuron integrates the messages it has received from other neurons and decides whether to fire an action potential.
axon terminal: buds located on the ends of axon branches that form synapses and release neurochemicals to other neurons.
axonal varicosity: a swelling in an axon wall that releases neurotransmitters through the wall via volume transmission.
axoplasmic transport: the movement of molecules in both directions along the outer surface of protein bundles called microtubules.
basal dendrite: a dendrite that horizontally branches out from the 30 μm base of the pyramid through the layer where the neuron resides.
basal ganglia: forebrain structures consisting of an egg-shaped nucleus that contains the putamen and globus pallidus and a tail-shaped structure called the caudate, which together are responsible for the production of movement. The basal ganglia have also been implicated in obsessive-compulsive disorder, Parkinson's disease, and Huntington's chorea.
beta rhythm: 12-38-Hz activity associated with arousal and attention generated by brainstem mesencephalic reticular stimulation that depolarizes neurons in both the thalamus and cortex. The beta rhythm can be divided into multiple ranges: beta 1 (12-15 Hz), beta 2 (15-18 Hz), beta 3 (18-25 Hz), and beta 4 (25-38 Hz).
bilateral synchronous slow waves: a pathological sign observed in drowsy children. When detected in alert adults, intermittent bursts of high-amplitude slow waves may signify gray matter lesions in deep midline structures.
brain-derived neurotrophic factor (BDNF): a growth-promoting protein that supports neuron survival and synapse formation and is essential for the late, lasting phase of long-term potentiation.
cation: positive ion, for example, sodium (Na+).
caudal: away from the front of the head.
cell body or soma: part of a neuron that contains machinery for cell life processes and receives and integrates EPSPs and IPSPs from axons generated by axosomatic synapses (junctions between axons and somas). The cell body of a typical neuron is 20 μm in diameter, and its spherical nucleus, which contains chromosomes comprised of DNA, is 5-10 μm across.
central nucleus of the amygdala: nucleus that orchestrates the nervous system's response to important stimuli by activating circuits in the brainstem (autonomic arousal) and the basal ganglia and periaqueductal gray (defensive behavior).
cerebral cortex: the layer of gray matter that covers the cerebral hemispheres. The cerebral cortex consists of gray matter and white matter.
chemical synapses: junctions between neurons that transmit molecules across a 20-50-nm (200-500-angstrom) synaptic cleft. Neurons use chemical synapses to produce short-duration (millisecond) and long-duration (seconds to days) changes in the nervous system. Chemical synapses are capable of more extensive communication and initiating more diverse and long-lasting changes than electrical synapses.
classical routes for EEG activation: specific sensory pathways like the visual (retina to the visual cortex), auditory (cochlea to the auditory cortex), and somatosensory (chemoreceptors and mechanoreceptors to the somatosensory cortex) systems. Increased transmission of information through these pathways desynchronizes EEG activity in the cortical regions to which these afferent neurons project, as specialized circuits of neurons independently process this information.
commissures: axon tracts. The left and right hemispheres communicate using the corpus callosum, anterior commissure, and posterior commissure.
complex: a sequence of waves.
COMT: a degrading enzyme that only targets the catecholamines dopamine and norepinephrine.
contingent negative variation (CNV): a steady, negative shift in potential (15 microvolts in young adults) detected at the vertex. This slow cortical potential may reflect expectancy, motivation, intention to act, or attention. The CNV appears 200-400 ms after a warning signal (S1), peaks within 400-900 ms when the S1-S2 interval is short, and sharply declines after a second stimulus that requires the performance of a response (S2).
continuous irregular delta: slow waves produced by white matter lesions seen in disorders like multiple sclerosis.
contralateral: structures that are located on opposite sides of the body. For example, neurons in the left primary motor cortex control muscles on the right side of the body.
corpus callosum: the largest commissure that connects the left and right frontal, parietal, and occipital lobes.
cortical excitability: the readiness of cortical neurons to generate action potentials in response to input. Neurofeedback can durably raise or lower it.
cortical negativity: a state where the cortical surface exhibits a negative electrical potential.
cortical neurons: nerve cells in the cortex responsible for generating and transmitting electrical impulses.
cortical positivity: a state where the cortical surface exhibits a positive electrical potential.
corticothalamic network: a unified network that generates diverse types of brain rhythms grouped by slow cortical oscillations.
cyclic AMP: a second messenger that moves about the neuron, activating other enzymes. Protein kinase A, which controls the excitability of ion channels, is a crucial enzyme target of cyclic AMP. Cyclic AMP also travels to the nucleus, where it can regulate gene expression.
D-serine: a neurotransmitter that binds to the glycine site on the NMDA receptor to trigger calcium entry into a dendritic spine when glutamate binds to its site, resulting in a large, prolonged increase in intracellular calcium.
Dale's law (also called Dale's principle): the superseded view that a neuron can release only one neurotransmitter. Many neurons in fact release more than one.
delta rhythm: 1-4-Hz oscillations generated by thalamocortical neurons and prominent during stage 3 (N3) sleep.
dendrite: a branched structure designed to receive messages from other neurons via axodendritic synapses (junctions between axons and dendrites), determining whether the axon hillock will initiate an action potential.
dendritic branch: a single segment or limb within that larger tree. It is one offshoot of a dendrite, often extending from a branch point (where a parent dendrite divides into daughter segments) to the next branch point or to a terminal tip. The dendritic tree is composed of many such branches.
dendritic spines: protrusions on the dendrite shaft where axons typically form axodendritic synapses.
dendritic tree: the complete, branching arrangement of dendrites that extends from a neuron's cell body (soma). It is the full collection of all the neuron's dendrites taken together, named for its tree-like, arborized shape. This structure is the neuron's main receptive surface, gathering synaptic inputs from other neurons and channeling those signals toward the soma.
dendrodendritic synapses: junctions between dendrites that communicate chemically across synapses and electrically across gap junctions.
depolarization: to make the membrane potential less negative by making the inside of the neuron less negative with respect to its outside.
diffusion: the distribution of molecules from areas of high concentration to low concentration.
diphasic wave: a wave that contains both a negative and positive deflection from the baseline.
dipole: the electrical field generated between the sink (where current enters the neuron ) and the source (place at the other end of the neuron where current leaves), which may be located anywhere along the dendrite.
dominant frequency: the EEG frequency with the most significant amplitude.
dopamine: a monoamine neurotransmitter exerts its postsynaptic effects on at least six receptors linked to G proteins. This means that dopamine functions as a neuromodulator. The two families include D1 (D1 and D5) and D2 (D2A, D2B, D3, and D4).
dorsal: toward the upper back or head.
dorsolateral prefrontal cortex: the left dorsolateral prefrontal cortex is concerned with approach behavior and positive affect. It helps us select positive goals and organizes and implements behavior to achieve these goals. The right dorsolateral prefrontal cortex organizes withdrawal-related behavior and negative affect and mediates threat-related vigilance. It plays a role in working memory for object location.
dual-action antidepressants: medications that activate 5-HT1 receptors to produce antidepressant and anxiolytic effects, while they blockade 5-HT2 (agitation, restlessness, and sexual dysfunction) and 5-HT3 (nausea, headache, and vomiting) receptors to minimize their side effects.
EEG activity: single wave or successive waves.
EEG power: the power in the EEG spectrum, proportional to the square of amplitude and expressed in µV2. Most EEG power falls within the 0-20 Hz frequency range. µV2 equals picowatts only if a 1-ohm reference resistance is assumed, and that assumption must be stated.
efferent: motoneuron that transmits information towards the periphery.
electrical synapse: symmetrical synapse where neurons communicate information bidirectionally across gap junctions between adjacent membranes using ions. Transmission across electrical synapses is nearly instantaneous, compared with the roughly 0.5-1-ms synaptic delay at chemical synapses. The rapid information transmission that characterizes electrical synapses enables large circuits of distant neurons to synchronize their activity and simultaneously fire.
electroencephalogram (EEG): the voltage difference between at least two electrodes, where at least one electrode is located on the scalp or inside the brain. The EEG is a recording of EPSPs and IPSPs that occur primarily in dendrites in pyramidal cells located in macrocolumns, several millimeters in diameter, in the upper cortical layers.
electrostatic pressure: the attractive or repulsive force between ions that moves them from one region to another.
entorhinal cortex: a structure located in the caudal region of the temporal lobe and receives pre-processed sensory information from all modalities and reports on cognitive operations. The entorhinal cortex provides the main input to the hippocampus, and is involved in memory consolidation, spatial localization, and provides input into the septohippocampal system that may generate the 4-7 Hz theta rhythm.
enzymatic deactivation: the process in which an enzyme breaks a neurotransmitter apart into inactive fragments. For example, acetylcholine transmission is ended by the enzyme acetylcholine esterase (AChE). Deactivating enzymes located in the synaptic cleft degrade a neurotransmitter molecule when it detaches from its binding site.
equilibrium potential: the membrane voltage at which the electrical and chemical forces acting on a particular ion are balanced, so there is no net flow of that ion across the membrane.
evoked potential: an event-related potential (ERP) elicited by external sensory stimuli (auditory, olfactory, somatosensory, and visual). An auditory evoked potential has a negative peak near 80-100 ms and a positive peak near 170 ms following stimulus onset; the pattern-reversal visual evoked potential instead peaks positively near 100 ms. The orienting response ("What is it?") is a sensory ERP. The N1-P2 complex in the auditory cortex of the temporal cortex reveals whether an uncommunicative person can hear a stimulus.
excitability: the ability of neurons to respond to stimuli and generate action potentials.
excitatory postsynaptic potential (EPSP): a brief positive shift in a postsynaptic neuron's potential produced when neurotransmitters bind to receptors and cause positive sodium ions to enter the cell. An EPSP pushes the neuron towards the threshold of excitation when it can initiate an action potential.
exocytosis: the process of neurotransmitter release. When an action potential arrives and depolarizes the terminal button, calcium ions enter the terminal button from the extracellular fluid. Calcium binds with clusters of protein molecules that connect the vesicles to the presynaptic membrane. The clusters move apart, forming a hole through both membranes called a fusion pore, and the neurotransmitter leaves the terminal button for the synaptic cleft or extracellular fluid.
exogenous ERP: an event-related potential (ERP) elicited by external sensory stimuli (auditory, olfactory, somatosensory, and visual).
explicit learning: behavioral changes that occur with our conscious awareness that require processing by the hippocampus.
extracellular dipole layers: macrocolumns of pyramidal cells, which lie parallel to the surface of the cortex, send opposite charges towards the surface and towards the deepest of the six layers of cortical neurons.
extracellular fluid: the fluid surrounding a neuron.
fast cortical potentials: EEG rhythms that range from 0.5 Hz-100 Hz. The main frequency ranges include delta, theta, alpha, sensorimotor rhythm, and beta.
feature binding: the process of linking information to perceptual objects (linking an apple's color to its shape) that may involve the 40-Hz rhythm.
fissures: deep grooves, for example, the lateral fissure.
focal waves: EEG waves that are detected within a limited area of the scalp, cerebral cortex, or brain.
frequency: the number of cycles completed each second expressed in hertz (Hz).
frequency synchrony: when identical EEG frequencies are detected at two or more electrode sites. For example, 12 Hz may be simultaneously detected at O1-A1 and O2-A2.
frontal lobes: the most anterior cortical lobes of the brain that are divided into the motor cortex, premotor cortex, and prefrontal cortex.
fusion pore: a hole through a vesicle and presynaptic membrane that allows neurotransmitter to leave the terminal button for the synaptic cleft or extracellular fluid.
G protein: a protein located inside a neuron's membrane next to a metabotropic receptor that is activated when the receptor binds a ligand. An alpha-subunit of the G protein then breaks away to perform actions within the cell.
GABA: an amino acid that is often inhibitory and that may be the most important inhibitory neurotransmitter in the brain. There are several types of GABA receptors, each of which produces inhibition differently.
gamma rhythm: EEG activity above about 30 Hz. Frequencies from 30-70 Hz are often called low gamma, while those above 70 Hz represent high gamma. These boundaries vary by author.
gap junction: narrow space between two cells bridged by connexons (protein channels) that allow ions to travel between them rapidly.
generalized asynchronous slow waves: waves that are seen in sleepy children and those with elevated temperatures. These waves may indicate degenerative disease, dementia, encephalopathy, head injury, high fever, migraine, and Parkinson's disease in adults.
glial cells: nonneural cells that guide, insulate, and repair neurons and provide structural, nutritional, and information-processing support. Glial cells contribute to slow cortical potentials (SCPs). Glial cells include astrocytes, microglia, oligodendrocytes, radial glial cells, and Schwann cells.
glutamate: an amino acid that is often excitatory and that may be the primary excitatory neurotransmitter in the brain. Its receptors are found on the surface of almost all neurons. There are at least 13 different receptors for glutamate, 5 ionotropic and 8 metabotropic. Most presynaptic neurons in the brain excite postsynaptic neurons via ionotropic glutamate receptors in the postsynaptic membrane. Metabotropic glutamate receptors may play a regulatory function, either augmenting or suppressing the activation of ionotropic glutamate receptors.
glycine: an amino acid that is often inhibitory and has a binding site on the NMDA receptor.
gray matter: brain tissue that looks grayish brown and comprises cell bodies, dendrites, unmyelinated axons, glial cells, and capillaries.
gyrus: ridge of cortex demarcated by sulci or fissures, for example, the precentral gyrus.
Hebbian plasticity: the strengthening of a synapse when its presynaptic and postsynaptic neurons are repeatedly active together, often summarized as neurons that fire together wire together.
hertz (Hz): the unit of frequency, an abbreviation for cycles per second.
hippocampus: a limbic structure located in the medial temporal lobe involved in 4-7 Hz theta activity, control of the endocrine system's response to stressors, formation of explicit memories, and navigation. Cortisol binding to this structure disrupts these functions, interferes with creating new neurons, and harms and kills hippocampal neurons.
hubs: highly centralized nodes through which other node pairs communicate; hubs allow efficient communication.
hyperpolarization: an increase in membrane potential, making the inside of a cell more negative relative to the outside.
immediate early genes: genes such as Arc that are transcribed within minutes of neural activity and whose protein products convert short-lived synaptic changes into lasting ones.
inhibitory postsynaptic potential (IPSP): a brief negative shift in a postsynaptic neuron's potential produced when cations like potassium leave a neuron or anions (negative ions) like chloride enter a neuron, which hyperpolarize the cell. An IPSP pushes the neuron away from its excitation threshold.
integration: the addition of EPSPs and IPSPs at the axon hillock. Neurons sum EPSPs and IPSPs over their surface in spatial integration and over milliseconds in temporal integration to raise the membrane from its resting potential to the excitation threshold. EPSPs and IPSPs last from 15-200 ms, while action potentials occur in 1-2 ms.
interneurons: neurons that receive input from and distribute output to other neurons. They typically have short processes and are largely, though not exclusively, confined to the central nervous system. They provide the integration required for decisions, learning and memory, perception, planning, and movement.
intracellular fluid: the watery cytoplasm contained within a neuron.
ion: a charged atom or molecule with a positive or negative charge. Positive ions are called cations, and negative ions are called anions.
ion concentration gradient: the unequal distribution of an ion across the neuronal membrane, such as sodium concentrated outside and potassium concentrated inside, which stores the energy that drives neural signaling.
ionotropic receptor: receptor protein that contains a binding site for a ligand and an ion channel that opens when the neurotransmitter attaches to this site.
ipsilateral: structures that are located on the same side of the body. For example, the left olfactory bulb distributes axons to the left hemisphere.
irregular waves: successive waves that constantly alter their shape and duration.
kappa rhythm: bursts of alpha or theta and is detected over the temporal lobes of subjects during cognitive activity.
lambda waves: saw-toothed transient waves from 20-50 μV in amplitude and 100-250 ms in duration detected over the occipital cortex during wakefulness. These positive deflections are time-locked to saccadic movements and observed during visual scanning, as during reading.
lateral: to the side, away from the center, as in the lateral geniculate nucleus.
lateral nucleus of the amygdala: a nucleus that processes sensory information and distributes it throughout the amygdala.
lateralized waves: waves that are primarily detected on one side of the scalp and that may indicate pathology.
Layer III: the cortical layer that is the primary source of corticocortical efferent fibers.
Layer IV: the cortical layer that is the primary destination of thalamocortical afferents and intra-hemispheric corticocortical afferents.
Layer V: the cortical layer that is the primary origin of efferent fibers that target subcortical structures that have motor functions.
Layer VI: the cortical layer that projects corticothalamic efferent fibers to the thalamus, which, together with the thalamocortical afferents, creates a dynamic and reciprocal relationship between these two structures.
Layers I-III: cortical layers that receive corticocortical afferent fibers that connect the left and right hemispheres.
left dorsolateral prefrontal cortex: the division of the prefrontal cortex concerned with approach behavior and positive affect. It helps us select positive goals and organizes and implements behavior to achieve these goals.
local field potential: the aggregate effect of the firing of the interconnected pyramidal neurons within the cortical columns plus additional mechanisms like glial cell modulation of the cortical electrical gradient.
local synchrony: synchrony that occurs when high-amplitude EEG signals are produced by the coordinated firing of cortical neurons.
localized slow waves: waves that may indicate a transient ischemic attack (TIA) or stroke, migraine, mild head injury, or tumors above the tentorium. Deep lesions result in bilateral or unilateral delta.
locus coeruleus: the noradrenergic branch of the ascending reticular activating system, which is responsible for vigilance. Subnormal norepinephrine transmission may contribute to ADHD.
long-latency potentials: potentials that have extended latencies following stimulus onset, for example, P300 and N400 ERPs.
long-term depression (LTD): a persistent decrease in synaptic strength following low-frequency stimulation.
long-term potentiation (LTP): a persistent increase in synaptic strength following high-frequency stimulation.
macrocolumns: circuits of cortical pyramidal neurons several millimeters in diameter that create extracellular dipole layers parallel to the surface of the cortex that send opposite charges towards the surface and towards the deepest of the six layers of cortical neurons. Since the pyramidal neurons are all aligned with the cortical surface, the postsynaptic potentials at cells within the same macrocolumn add together. This summation occurs because they share the same charge and the macrocolumns fire synchronously.
medial: toward the center of the body, away from the side. For example, the medial geniculate nucleus.
medial prefrontal cortex: the division of the prefrontal cortex that integrates cognitive-affective information and helps control the hypothalamic-pituitary-adrenal (HPA) axis during emotional stress.
membrane potential: a neuron's electrical charge created by a difference in ion distribution within and outside the neuron. A typical resting potential is about -70 mV (thousandths of a volt), since the inside of a resting axon is more negatively charged than the outside.
mesocortical neurons: dopaminergic neurons that project from the ventral tegmental area of the midbrain to the prefrontal cortex and excite prefrontal cortical neurons that control working memory, planning, and strategy preparation for problem-solving. Underactivity in this pathway is associated with the negative symptoms of schizophrenia-like attentional deficits.
metabotropic receptors: include all G protein-linked receptors located on neurons, including autoreceptors. Neurotransmitters that bind to G protein-linked receptors are often called neuromodulators. Metabotropic receptors, which indirectly control the cell's operations, expend energy, and produce slower, longer lasting, and more diverse changes than ionotropic receptors. Their effects can last several seconds, instead of milliseconds, because of the long-lived activity of G proteins and cyclic AMP.
microglia: microscopic glial cells that participate in the immune response.
microtubules: hollow cylindrical protein bundles that are involved in axoplasmic transport.
modulating effects: neuromodulators like the monoamines alter the performance of diffuse networks of target neurons by indirectly controlling cellular operations when they bind to metabotropic receptors.
module: a set of interconnected nodes in a neural network.
monoamine neurotransmitters: amine neurotransmitters that include dopamine, norepinephrine, epinephrine (catecholamines), and serotonin (indoleamine). These neurotransmitters are released using volume transmission and generally have modulating effects, altering the performance of diffuse networks of target neurons.
monoamine oxidase (MAO): an enzyme that degrades and inactivates the monoamine neurotransmitters dopamine, norepinephrine, and serotonin.
monoamine oxidase inhibitors (MAOIs): antidepressant drugs that interfere with MAO's breakdown of monoamines and increase monoamine availability to treat clinical depression.
monophasic wave: either a single negative (upward) or positive (downward) deflection from baseline.
motor cortex: the subdivision of the frontal lobe located in the precentral gyrus and guides fine motor coordination (like writing).
motor ERPs: event-related potentials detected over the primary motor cortex (precentral gyrus) during movement. Their amplitude is proportional to the force and rate of skeletal muscle contraction.
motor neurons: efferent neurons that convey commands to glands, muscles, and other neurons.
movement-related potentials (MRPs): slow cortical potentials that occur at 1 second as subjects prepare for unilateral voluntary movements. MRPs are distributed bilaterally with maximum amplitude at Cz. The supplementary motor area and primary motor and somatosensory cortices primarily generate these potentials.
mu rhythm: 7-11-Hz waves resemble wickets and appear as several-second trains over central or centroparietal sites (C3 and C4).
multiple spike-and-slow-wave complex: multiple spikes associated with at least one slow wave.
muscarinic receptors: metabotropic ACh receptors that are stimulated by muscarine and blocked by atropine. Muscarinic receptors control smooth muscle and predominate in the CNS. In the CNS, muscarinic receptors help mediate learning, memory, attention, arousal, EEG, and postural control.
myelinated axons: axons that are insulated by myelin by oligodendrocytes in the central nervous system and Schwann cells in the peripheral nervous system.
N1-P2: a sensory event-related potential in the auditory cortex of the temporal cortex that reveals whether an uncommunicative person can hear a stimulus.
N400 potential: an event-related potential (ERP) elicited when we encounter semantic violations like ending a sentence with a semantically incongruent word ("The handsome prince married the beautiful fish"), or when the second word of a pair is unrelated to the first (BATTLE/GIRL).
negative SCPs: slow cortical potentials associated with depolarization of the apical dendrites of cortical pyramidal cells, with a contribution from glia, that increase the probability of neuron firing.
neuroaxis: an imaginary line that runs centrally through the central nervous system (CNS) from the front of the prefrontal cortex to the base of the spinal cord.
neuromodulator: a neurochemical that modifies the effect of neurotransmitters through mechanisms like binding to metabotropic receptors.
neuron: a nerve cell that is the fundamental anatomical unit of the nervous system.
neuroplasticity: the remodeling of neurons and neural networks with experience, the foundational mechanism that makes neurofeedback learning possible.
nicotinic ACh receptor: an ionotropic receptor that is stimulated by nicotine and blocked by curare. They are mainly found in the PNS on skeletal muscles. At CNS axoaxonic synapses, they produce presynaptic facilitation (increase neurotransmitter release). In the CNS, nicotinic receptors help regulate cortical blood flow, anxiety reduction, and decision-making.
nigrostriatal pathway: dopaminergic pathway from the substantia nigra to the basal ganglia (caudate nucleus and putamen) that controls movement. The nigrostriatal pathway is progressively destroyed in Parkinson's disease.
nitric oxide: a gaseous retrograde transmitter that is involved in long-term potentiation (LTP).
NMDA (glutamate) receptors: ligand-gated and voltage-gated glutamate receptors that bind the glutamate agonist NMDA. NMDA receptors play an essential role in long-term potentiation (LTP).
node: vertex within a neural network.
nodes of Ranvier: gaps between myelinated axon segments where the axon membrane is exposed to extracellular fluid and action potentials are regenerated by sodium ion entry.
norepinephrine: a monoamine neurotransmitter that exerts postsynaptic effects at alpha and beta receptors, each with two subtypes. All norepinephrine receptors are G protein-linked. The cell bodies of the core noradrenergic system are located in the locus coeruleus, a nucleus found in the dorsal pons.
nucleus accumbens: a limbic structure that is a target of dopamine released by the mesolimbic pathway. The nucleus accumbens plays a critical role in the reinforcement of diverse activities, including ingestion of drugs like central nervous system stimulants.
Nyquist theorem: the principle that a signal must be sampled at least twice its highest frequency of interest to be represented accurately. Sampling too slowly causes aliasing, in which fast activity appears as a slower, false rhythm.
occipital lobes: cortical lobes that are posterior to the parietal lobes. They process visual information from the eyes in collaboration with the frontal, parietal, and temporal lobes.
odd-ball stimulus: a meaningful stimulus that is different from others in a series used to elicit the P300 potential. For example, a colored playing card in a series of monochrome cards.
oligodendrocytes: glial cells that insulate adjacent axons within the brain and spinal cord of the central nervous system.
orbitofrontal cortex: the frontal lobe subdivision that is concerned with affective evaluation. It decodes the punishment and reward value of stimuli and helps inhibit inappropriate behavior. Phineas Gage's profound personality changes were produced by damage to this region.
orienting response: Pavlov's "What is it?" reaction to stimuli like the sound of a vase crashing that includes (1) increased sensory sensitivity, (2) head (and ear) turning toward the stimulus, (3) increased muscle tone (reduced movement), (4) EEG desynchrony, (5) peripheral constriction and cephalic vasodilation, (6) a rise in skin conductance, (7) heart rate slowing, and (8) slower, deeper breathing.
P300 potential: an event-related potential (ERP) whose peak latency typically falls between 250 and 500 ms, lengthening with task difficulty, age, and cognitive impairment. The largest amplitude positive peaks are located over the parietal lobe. The P300 potential may reflect an event's subjective probability, meaning, and transmission of information.
paradoxical negativity: in the context of SCPs, it refers to surface-negative EEG shifts when neurons are depolarized due to volume conduction of negative potentials from the extracellular space to the scalp.
paradoxical positivity: in the context of SCPs, it refers to surface-positive EEG shifts when neurons are hyperpolarized due to volume conduction of positive potentials from the extracellular space to the scalp.
paralimbic cortex: a transitional region between neocortex and allocortex.
parietal lobes: cortical lobes posterior to the frontal lobes that are divided into the primary somatosensory cortex (postcentral gyrus) and secondary somatosensory cortex. Their primary function is to process somatosensory information like pain and touch. The right posterior parietal lobe helps guide movements, locate objects in three-dimensional space, and create body boundaries.
Parkinson's disease (PD): a progressive neurodegenerative disorder characterized primarily by motor symptoms such as tremor, rigidity, bradykinesia (slowness of movement), and postural instability.
phase: the degree to which the peaks and valleys of EEG waveforms coincide.
phase synchrony: synchrony when identical EEG frequencies are detected at two or more electrode sites, and the peaks and valleys of the EEG waveforms coincide. This is also called global synchrony. For example, EEG training may produce phase-synchronous 12-Hz alpha waves at O1-A1 and O2-A2.
polarization: to make the membrane potential more negative by making the inside of the neuron more negative with respect to its outside.
polyphasic (multiphasic) wave: a wave that contains two or more deflections of opposite polarity from baseline.
positive SCPs: slow cortical potentials associated with hyperpolarization of the apical dendrites of cortical pyramidal cells, with a contribution from glia, that decrease the probability of neuron firing.
posterior: near or toward the back of the head.
posterior commissure: axon tracts located below the corpus callosum that connect the right and left diencephalon and mesencephalon.
precision: the number of voltage gradations or steps.
prefrontal cortex: the most anterior frontal lobe division and is subdivided into dorsolateral, medial, orbitofrontal, and anterior cingulate regions responsible for executive functions like attention and planning.
premotor cortex: the frontal lobe subdivision that is anterior to the motor cortex and helps program head, trunk, and limb movements.
presynaptic facilitation: a modulatory process in which a neuron increases the presynaptic neuron's neurotransmitter release by delivering a neurotransmitter that increases calcium ion entry into its terminal button.
presynaptic inhibition: a modulatory process in which a neuron decreases neurotransmitter release by reducing calcium ion entry.
primary somatosensory cortex (S1): the parietal lobe subdivision located at the postcentral gyrus that processes information about touch and pain.
protein kinase A: an intracellular enzyme that controls the excitability of ion channels and is a critical enzyme target of cyclic AMP.
radial dipole: a current dipole oriented perpendicular to the scalp, as in the crown of a gyrus. Because its field projects directly toward the electrodes, the scalp EEG detects it well.
raphe nuclei: serotonergic cell bodies in the midbrain, pons, and medulla give rise to most of the brain's serotonergic neurons.
rate law: the principle that neurons represent the intensity of a stimulus by variation in the rate of axon firing.
readiness potential: slow-rising, negative potential (10-15 µV) detected at the vertex before voluntary and spontaneous movement. Its late phase precedes voluntary movement by 0.5 to 1 second and peaks at about the time the subject responds; an earlier component begins roughly 1.5 to 2 seconds before movement onset.
regular or monomorphic waves: successive waves with identical shapes. Regular waves may resemble sine waves (sinusoidal) or maybe arched (resembling wickets) or saw-toothed (asymmetrical and triangular).
relative refractory period: the interval following the absolute refractory period when a stronger-than-usual input is needed to fire the neuron. It allows the rate law to encode stimulus intensity.
resting membrane potential: the steady voltage across a neuron's membrane when it is not signaling, typically about -70 mV with the inside negative relative to the outside. It is established by ion concentration gradients and selective permeability and maintained by the sodium-potassium pump.
resting potential: the membrane potential of a neuron when it is not influenced by messages from other neurons.
reticular formation: a network of 90 nuclei within the central brainstem from the lower medulla to the upper midbrain. The reticular formation sends axons to the spinal cord, thalamus, and cortex, contributing to diverse functions like neurological reflexes, muscle tone and movement, attention, arousal, and sleep.
reuptake: the primary method that neurons terminate the action of neurotransmitters. Reuptake transporters located in terminal buttons and astrocytes remove neurotransmitters from the synaptic cleft.
reward deficiency syndrome: Blum's hypothesis that an abnormal form of the A1 allele is present in most severe alcoholics and results in defective D2 receptors. Reduced D2 receptor activity may reduce the activation of the nucleus accumbens and hypothalamus and result in dysphoria, drug craving, and compulsive drug-seeking and abuse.
right dorsolateral prefrontal cortex: the division of the prefrontal cortex that organizes withdrawal-related behavior and negative affect and mediates threat-related vigilance. It plays a role in working memory for object location.
rostral: toward the front of the head.
saltatory conduction: action potential conduction in myelinated axons in which action potentials jump from node to node for 200 times greater speed.
sampling rate: the number of measurements per second (Hz).
Schwann cells: glial cells that provide myelin for single PNS axons and facilitate axonal regeneration following damage.
secondary somatosensory cortex (S2): a region of the parietal lobe that receives somatosensory information from the primary somatosensory cortex (S1).
selective permeability: the property of the neuronal membrane that allows some ions to cross more readily than others. At rest, the membrane is far more permeable to potassium than to sodium.
sensorimotor rhythm (SMR): EEG rhythm that ranges from 12-15 Hz and is located over the sensorimotor cortex (central sulcus). The waves are synchronous. The sensorimotor rhythm is associated with the inhibition of movement and reduced muscle tone. The SMR is generated by "ventrobasal relay cells in the thalamus and thalamocortical feedback loops."
sensorimotor system: in Sterman's model, ascending pathways that convey information about touch and proprioception to the thalamus, the thalamus and its thalamic projections to the sensorimotor cortex, and the sensorimotor cortex, and its efferent fibers.
sensory event-related potentials (ERPs): event-related potentials evoked by external sensory stimuli (auditory, olfactory, somatosensory, and visual). In the auditory modality these evoked potentials or exogenous ERPs have a negative peak near 80-100 ms and a positive peak near 170 ms following stimulus onset. These changes in brain activity in response to specific stimuli. ERPs can be detected throughout the cortex. Investigators monitor ERPs by placing electrodes at locations like the midline (Fz, Cz, and Pz). A computer analyzes a subject's EEG responses to the same stimulus or task over many trials to subtract random EEG activity. For a fixed stimulus, task, and electrode site the averaged ERP is highly replicable, with peaks at characteristic latencies; morphology and scalp topography differ across modalities and tasks.
sensory neurons: neurons specialized for sensory intake. They are called afferent because they transmit sensory information towards the central nervous system (brain and spinal cord).
septal nuclei: in Sieb's model, when the prefrontal cortex receives information about high-priority environmental events, it signals cell bodies in the septum to induce a beta rhythm in the hippocampus to remove its inhibition of vigilance centers.
septohippocampal system: a subcortical circuit from the septum to hippocampus that contributes to 4-7 Hz theta activity.
septum: a limbic structure that contains several nuclei involved in emotion and addiction and control of aggressive behavior.
sharp transients: a sequence that contains several sharp waves.
sharp waves: waves that resemble spikes with a pointed peak with a longer 70-200-ms duration.
sink: a site where current enters the neuron. Positive sodium ion entry into a neuron creates an active sink, represented by -ve.
slow cortical potentials (SCPs): gradual changes in the membrane potentials of cortical dendrites that last from 300 ms to several seconds. These potentials include the contingent negative variation (CNV), the readiness potential, and movement-related potentials (MRPs). SCPs are the low-frequency subclass of event-related potentials; the faster P300 and N400 components are event-related potentials but not slow cortical potentials. SCPs modulate the firing rate of cortical pyramidal neurons by exciting or inhibiting their apical dendrites. They group the classical EEG rhythms using these synchronizing mechanisms.
sodium (Na+) ions: positive ions that enter a neuron during EPSPs and action potentials.
sodium-potassium pump: the ATP-powered transporter that exchanges three sodium ions moved out of the neuron for every two potassium ions moved in, maintaining the ion gradients that produce the resting membrane potential. Also called the sodium-potassium transporter.
sodium-potassium transporters: pumps that are powered by ATP and that exchange three sodium for two potassium ions.
soma or cell body: the region of a neuron that contains machinery for cell life processes and receives and integrates EPSPs and IPSPs from axons generated by axosomatic synapses (junctions between axons and somas). The cell body of a typical neuron is 20 μm in diameter, and its spherical nucleus, which contains chromosomes comprised of DNA, is 5-10 μm across.
source: the place at the end of the neuron opposite of the sink where current leaves, represented by +ve. The extracellular area surrounding the source becomes electrically positive.
spatial summation: the addition of EPSPs and IPSPs over a neuron's surface.
spike: a negative transient with a pointed peak at conventional paper speeds, 20-70-ms duration, and 40-100 μV amplitude; rare dendritic action potentials.
spike-and-slow-wave complex: a spike followed by a higher amplitude slow wave at 3 Hz. In an absence seizure, the amplitudes are very high (e.g., 160 μV).
spindle behavior: the waxing-and-waning, fusiform amplitude envelope of a transient oscillatory burst in the EEG: the rhythm's amplitude progressively grows to a peak and then tapers off, rather than holding a constant level, producing the spindle-like (thread-on-a-spindle) shape on the trace.
spindle waves: waves that originate in the thalamus and occur during unconsciousness and stage II sleep.
spiny neurons: neurons with dendritic spines that are usually excitatory.
stimulus-preceding negativity (SPN): a slow negative potential shift observed before a stimulus that signals important or relevant information, such as feedback or a reward. SPN reflects anticipatory attention and affective processes involving regions like the insula and orbitofrontal cortex. SPN is associated with emotional and cognitive anticipation.
striatal: basal ganglia (caudate nucleus and putamen).
Stroop test: cognitive monitoring task where color and names conflict.
substantia nigra: midbrain structure that projects to the basal ganglia (caudate nucleus and putamen) to control movement and that is progressively destroyed in Parkinson's disease.
sulcus: a shallow groove in the surface of the cerebral hemisphere, for example, the central sulcus.
surface-negative: a negative SCP shift typically associated with increased cortical excitability and response readiness.
surface-positive: a positive SCP shift typically associated with decreased cortical excitability and relaxation.
synapse-associated polyribosome complexes (SPRCs): organelles with dendrites that can produce proteins that allow rapid remodeling of synapses. A polyribosome complex consists of several ribosomes bound to messenger RNA (mRNA). SPRCs represent one mechanism underlying synaptic plasticity.
synaptic cleft: 20-50-nm fluid-filled gap between presynaptic and postsynaptic structures.
synaptogenesis: the formation of new synapses between neurons.
synchronous: adverb meaning that groups of neurons depolarize and hyperpolarize simultaneously.
synchrony: the coordinated firing of pools of neurons. EEG signals can display local synchrony, frequency synchrony, and phase synchrony.
tangential dipole: a current dipole oriented parallel to the scalp, as in the wall of a sulcus. Much of its field runs sideways beneath the electrodes, and opposing sulcal walls can cancel, so the scalp EEG detects it poorly.
telencephalon: the frontal subdivision of the forebrain, including the cerebral cortex, basal ganglia, and limbic system.
temporal lobes: lobes separated from the rest of the cortical lobes by the Sylvian fissure. The temporal lobes process hearing, smell, and taste information and help us understand spoken language and recognize visual objects and faces. The amygdala and hippocampus, which lie beneath the temporal cortex, play crucial roles in emotion, declarative, emotional, and working memory, and navigation.
temporal summation: the addition of EPSPs and IPSPs over time. Summation is more effective when postsynaptic potentials are generated more closely in time.
terminal buttons: buds located on the ends of axon branches that form synapses and release neurochemicals to other neurons. They contain vesicles that store neurotransmitters for release when an action potential arrives. A terminal button's presynaptic membrane may possess reuptake transporters that return neurotransmitters from the synapse or extracellular space for repackaging.
thalamus: forebrain structure above the hypothalamus that receives, filters, and distributes most sensory information. The thalamus contains neurons that can block or relay ascending sensory information. When these thalamic neurons rhythmically fire, this blocks the transmission of information to the cortex. When they depolarize in response to sensory information, this integrates and transmits this information to the cortex. Inputs to the thalamus determine whether these neurons block or relay sensory information.
theta rhythm: 4-7-Hz rhythms, which some authors extend to 8 Hz, generated by a cholinergic septohippocampal system that receives input from the ascending reticular formation and a noncholinergic system that originates in the entorhinal cortex, which corresponds to Brodmann areas 28 and 34 at the caudal region of the temporal lobe.
threshold of excitation: the membrane potential at which an axon initiates an action potential, nominally -55 mV.
transient: a single wave or sequence of regular waves, called a complex, distinguishable from background EEG activity.
traveling waves: EEG oscillations that move across the cortex that may mediate large-scale coordination of brain networks and support connectivity.
triphasic wave: a wave that contains three deflections from baseline.
unmyelinated axons: smaller-diameter axons without fatty insulation that conduct more slowly than myelinated axons.
ventral: toward the base of the skull or front of the body.
ventral striatum: the olfactory tubercle and nucleus accumbens.
ventral tegmental area: the midbrain structure that distributes dopaminergic axons to the nucleus accumbens. Serotonin receptors on endorphin-releasing neurons in the hypothalamus may increase the activity of dopaminergic reward pathways by inhibiting the release of GABA at receptors on cell bodies of the ventral tegmental area neurons.
vigilance system: in Sterman's model, a system that consists of both specific brainstem nuclei (e.g., locus coeruleus and raphe nuclei) and their diffuse connections with the thalamus and other subcortical structures, and the cortex. Several neurotransmitter systems mediate vigilance, including cholinergic/glutamatergic (reticular formation), noradrenergic (locus coeruleus), and serotonergic (raphe) neurons.
voltage-gated ion channel: a membrane channel that opens or closes in response to changes in membrane voltage. Voltage-gated sodium and potassium channels generate the action potential.
volume conduction: the passive spread of electrical current from a neural source through brain, cerebrospinal fluid, skull, and scalp. It smears each source across the scalp, limiting the spatial resolution of the EEG. It is distinct from volume transmission.
volume transmission: extrasynaptic neurotransmitter release from axonal varicosities, dendrites, and terminal buttons into the extracellular space. Monoamines like norepinephrine and serotonin are released outside the synaptic cleft.
waveform: the shape and form of an EEG signal.
white matter: the layer beneath the cortex that mainly consists of myelinated axons.
+ve: the source is the place at the other end of the neuron where current leaves, and is represented by +ve.
-ve: a sink is where current enters the neuron. Positive sodium ion entry into a neuron creates an active sink, represented by -ve.
Test Yourself on ClassMarker
Click on the ClassMarker logo below to take a 10-question exam over this entire unit. There is no password.

Review Flashcards on Quizlet
Click on the Quizlet logo to review our chapter flashcards.

Visit the BioSource Software Website
BioSource Software offers Physiological Psychology, which satisfies BCIA's Neuroanatomy requirement, and the Neurofeedback100 Testing Service, which provides extensive multiple-choice testing over the Neurofeedback Blueprint.

Assignment
Now that you have completed this unit, how would you explain the relationship between local field potentials and the EEG? How does anatomy explain why the EEG is comprised of EPSPs and IPSPs instead of action potentials?
References
Advokat, C. D., Comaty, J. E., & Julien, R. M. (2019). Julien's primer of drug action (15th ed.). Worth Publishers.
Aloisi, F. (2001). Immune function of microglia. Glia, 36, 165-179. https://doi.org/10.1002/glia.1106
Altenmuller, E. O., & Gerloff, C. (1999). Psychophysiology and the EEG. In E. Niedermeyer & F. Lopes da Silva (Eds.), Electroencephalography: Basic principles, clinical applications, and related fields (4th ed.). Williams and Wilkins.
Amzica, F., & Steriade, M. (2002). The functional significance of K-complexes. Sleep Medicine Reviews, 6(2), 139–149. https://doi.org/10.1053/smrv.2001.0181
Andreassi, J. L. (2007). Psychophysiology: Human behavior and physiological response (5th ed.). Lawrence Erlbaum and Associates, Inc.
Anjum, M., Smyth, C., Dijk, D., Starr, P., Denison, T., & Little, S. (2023). Multi-night cortico-basal recordings reveal mechanisms of NREM slow wave suppression and spontaneous awakenings at high-temporal resolution in Parkinson’s disease. Research Square. https://doi.org/10.21203/rs.3.rs-3484527/v1
Arnsten, A. F. (2006). Fundamentals of Attention-Deficit/Hyperactivity Disorder: Circuits and pathways. Journal of Clinical Psychiatry, 67 (Suppl. 8), 7-12.
Azevedo, F. A. C., Carvalho, L. R. B., Grinberg, L. T., Farfel, J. M., Ferretti, R. E. L., Leite, R. E. P., Jacob Filho, W., Lent, R., & Herculano-Houzel, S. (2009). Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. The Journal of Comparative Neurology, 513(5), 532-541. https://doi.org/10.1002/cne.21974
Babiloni, C., Babiloni, F., Carducci, F., Cincotti, F., Del Percio, C., Hallett, M., Moretti, D. V., Romani, G. L., & Rossini, P. M. (2002). High resolution EEG of sensorimotor brain functions: Mapping ERPs or mu ERD? In R. C. Reisin, M. R. Nuwer, M. Hallett, & C. Medina (Eds.). Advances in Clinical Neurophysiology (Supplements to Clinical Neurophysiology Vol. 54). Elsevier Science B. V.
Basile, L. F. H., Yacubian, J., Ferreira, B. L. C., Valim, A. C., & Gattaz, W. F. (2004). Topographic abnormality of slow cortical potentials in schizophrenia. Brazilian Journal of Medical and Biological Research, 37(1), 97-109. https://doi.org/10.1590/s0100-879x2004000100014
Bear, M. F., Connors, B. W., & Paradiso, M. A. (2020). Neuroscience: Exploring the brain (4th ed., enhanced ed.). Jones & Bartlett Learning.
Bear, M. F., Connors, B. W., & Paradiso, M. A. (2026). Neuroscience: Exploring the brain (5th ed.). Jones & Bartlett Learning.
Birbaumer, N. (1999). Slow cortical potentials: Plasticity, operant control, and behavioral effects. The Neuroscientist, 5, 74–78. https://doi.org/10.1177/107385849900500211
Birbaumer, N., Elbert, T., Canavan, A. G., & Rockstroh, B. (1990). Slow potentials of the cerebral cortex and behavior. Physiological Reviews, 70(1), 1–41. https://doi.org/10.1152/physrev.1990.70.1.1
Breedlove, S. M., & Watson, N. V. (2023). Behavioral neuroscience (10th ed.). Sinauer Associates, Inc.
Brienza, M., & Mecarelli, O. (2019). Neurophysiological basis of EEG. In O. Mecarelli (Ed.), Clinical electroencephalography (pp. 25-45). Springer. https://doi.org/10.1007/978-3-030-04573-9_2
Brini, M., Cali, T., Ottolini, D., & Carafoli, E. (2014). Neuronal calcium signaling: Function and dysfunction. Cellular and Molecular Life Sciences, 71(15), 2787–2814. https://doi.org/10.1007/s00018-013-1550-7
Brunia, C. H. M., van Boxtel, G. J. M., & Böcker, B. E. (2012). Negative slow waves as indices of anticipation: The Bereitschaftspotential, the Contingent Negative Variation, and the Stimulus-Preceding Negativity. In E. S. Kappenman & S. J. Luck (Eds.), The Oxford handbook of Event-Related Potential components (pp. 190-208). Oxford Academic. https://doi.org/10.1093/oxfordhb/9780195374148.013.0108
Bullmore, E., & Sporns, O. (2009). Complex brain networks: Graph theoretical analysis of structural and functional systems. Nature Reviews Neuroscience, 10, 186-198. https://doi.org/10.1038/nrn2575
Buzsáki, G. (2006). Rhythms of the brain. Oxford University Press.
Cameron, H. A., & Dayer, A. G. (2008). New interneurons in the adult neocortex: Small, sparse, but significant? Biological Psychiatry, 63(7), 650-655. https://dx.doi.org/10.1016%2Fj.biopsych.2007.09.023
Carlson, N. R., & Birkett, M. A. (2021). Physiology of behavior (13th ed.). Pearson.
Caton, R. (1875). The electric currents of the brain. British Medical Journal, 2, 278.
Chan, C. Y., Ke, D. S., & Chen, J. Y. (2009). Essential fatty acids and human brain. Acta Neurol Taiwan, 18(4), 231-241. PMID: 20329590
Collura, T. F. (2014). Technical foundations of neurofeedback. Taylor & Francis.
Costanzo, R. M. (1991). Regeneration of olfactory receptor cells. Ciba Foundation Symposium, 160, 233-242. https://doi.org/10.1002/9780470514122.ch12
Creutzfeldt, O. D. (1995). Cortex cerebri. Oxford University Press.
Damasio, A. (2010). Self comes to mind. Pantheon Books.
Daum, I., Rockstroh, B., Birbaumer, N., Elbert. T., Canavan, A., & Lutzenberger W. (1993). Behavioural treatment of slow cortical potentials in intractable epilepsy: Neuropsychological predictors of outcome. Journal of Neurology, Neurosurgery & Psychiatry, 56(1), 94-97. https://doi.org/10.1136/jnnp.56.1.94
deCharms, R. C., Fumiko, M., Glover, G. H., Ludlow, D., Pauly, J. M., Soneji, D., Gabrieli, J. D. E., & Mackey, S. C. (2005). Control over brain activation and pain learned by using real-time functional MRI. Proceedings of the National Academy of Sciences, 102(51), 18626-18631. https://doi.org/10.1073/pnas.0505210102
DeLong, M. R. (1990). Primate models of movement disorders of basal ganglia origin. Trends in Neurosciences, 13(7), 281-285. https://doi.org/10.1016/0166-2236(90)90110-v
Demos, J. N. (2019). Getting started with neurofeedback. (2nd ed.). W. W. Norton & Company.
di Pellegrino, G., Fadiga, L., Fogassi, L., Gallese, V., & Rizzolatti, G. (1992). Understanding motor events: A neurophysiological study. Experimental Brain Research, 91(1), 176-180. https://doi.org/10.1007/BF00230027
Eisenberger, N. I., Lieberman, M. D., & Williams, K. D. (2003). Does rejection hurt? An fMRI study of social exclusion. Science, 302, 290-292. https://doi.org/10.1126/science.1089134
El-Boustani, S., Ip, J., Breton-Provencher, V., Knott, G., Okuno, H., Bito, H., & Sur, M. (2018). Locally coordinated synaptic plasticity of visual cortex neurons in vivo. Science, 360(6395), 1349-1354. https://doi.org/10.1126/science.aao0862
Elbert, T., Rockstroh, B., Lutzenberger, W., & Birbaumer, N. (1980). Biofeedback of slow cortical potentials. I. Electroencephalography and Clinical Neurophysiology, 48(3), 293-301. https://doi.org/10.1016/0013-4694(80)90265-5
Enticott, P. G., Kennedy, H. A., Rinehart, N. J., Tonge, B. J., Bradshaw, J. L., Taffe, J. R., Daskalakis, Z. J., & Fitzgerald, P. B. (2012). Mirror neuron activity associated with social Impairments but not age in Autism Spectrum Disorder. Biological Psychiatry, 71(5), 427-433. https://doi.org/10.1016/j.biopsych.2011.09.001
Eriksson, P. S., Perfilieva, E., Björk-Eriksson, T., Alborn, A. M., Nordborg, C., Peterson, D. A., & Gage, F. H. (1998). Neurogenesis in the adult human hippocampus. Nature Medicine, 4(11), 1313-1317. https://doi.org/10.1038/3305
Evans, J. R., & Abarbanel, A. (1999). Introduction to quantitative EEG and neurofeedback. Academic Press.
Farwell, L. A., & Donchin, E. (1991). The truth will out: Interrogative polygraphy (“lie detection”) with event-related brain potentials. Psychophysiology, 28, 531–547. https://doi.org/10.1111/j.1469-8986.1991.tb01990.x
Fischer, F., Hamann, A., & Osiewacz, H. D. (2020). Mitochondrial quality control: An integrated network of pathways. Trends in Biochemical Sciences, 45(3), 179–190. https://doi.org/10.1016/j.tibs.2019.12.008
Fox, S. I., & Rompolski, K. (2022). Human physiology (16th ed.). McGraw-Hill.
Garrett, B. (2003). Brain and behavior. Thompson/Wadsworth.
Green, D. R., Galluzzi, L., & Kroemer, G. (2011). Mitochondria and the autophagy–inflammation–cell death axis in organismal aging. Science, 333(6046), 1109–1112. https://doi.org/10.1126/science.1201940
Hansson, E., & Rönnbäck, L. (2003). Glial neuronal signaling in the central nervous system. FASEB J, 17, 341-348. https://doi.org/10.1096/fj.02-0429rev
Harris, J. J., Jolivet, R., & Attwell, D. (2012). Synaptic energy use and supply. Neuron, 75(5), 762–777. https://doi.org/10.1016/j.neuron.2012.08.019
Heinrich, H., Gevensleben, H., Freisleder, F. J., Moll, G. H., & Rothenberger, A. (2004). Training of slow cortical potentials in attention-deficit/hyperactivity disorder: Evidence for positive behavioral and neurophysiologic effects. Biological Psychiatry, 55(7), 772–775. https://doi.org/10.1016/j.biopsych.2003.11.013
Hoedlmoser, K., Pecherstorfer, T., Gruber, G., Anderer, P., Doppelmayr, M., Klimesch, W., & Schabus, M. (2008). Instrumental conditioning of human sensorimotor rhythm (12-15 Hz) and its impact on sleep as well as declarative learning. Sleep, 31(10), 1401–1408.
Hugdahl, K. (1995). Psychophysiology: The mind-body perspective. Harvard University Press.
Kalat, J. W. (2019). Biological psychology (13th ed.). Cengage Learning.
Kalia, L. V., & Lang, A. E. (2015). Parkinson's disease. Lancet (London, England), 386(9996), 896–912. https://doi.org/10.1016/S0140-6736(14)61393-3
Kennerley, S. W., Behrens, T. E., & Wallis, J. D. (2011). Double dissociation of value computations in orbitofrontal and anterior cingulate neurons. Nat Neurosci, 14(12), 1581-1589. https://doi.org/10.1038/nn.2961
Kerson, C., Sherlin, L. H., & Davelaar, E. J. (2025). Neurofeedback, biofeedback, and basic learning theory: Revisiting the 2011 conceptual framework. Applied Psychophysiology and Biofeedback. Advance online publication. https://doi.org/10.1007/s10484-025-09756-4
Kitamura, T., Saitoh, Y., Takashima, N., Murayama, A., Niibori, A., Ageta, H., ... Inokuchi, K. (2009). Adult neurogenesis modulates the hippocampus-dependent period of associative fear memory. Cell, 139(4), 814-827. https://doi.org/10.1016/j.cell.2009.10.020
Klein, S. B., & Thorne, B. M. (2007). Biological psychology. Worth Publishers.
Kotchoubey, B., Blankenhorn, V., Fröscher, W., Strehl, U. & Birbaumer, N. (1997). Stability of cortical self-regulation in epilepsy patients. Neuroreport, 8(8), 1867-1870. https://doi.org/10.1097/00001756-199705260-00015
Kotchoubey, B., Busch, S., Strehl, U. & Birbaumer, N. (1999). Changes in EEG power spectra during biofeedback of slow cortical potentials in epilepsy. Applied Psychophysiology and Biofeedback, 24(4), 213-233. https://doi.org/10.1023/a:1022226412991
Kotchoubey, B., Kübler, A., Strehl, U., Flor, H., & Birbaumer, N. (2002). Can humans perceive their brain states? Consciousness and Cognition, 11(1), 98-113. https://doi.org/10.1006/ccog.2001.0535
Kotchoubey, B., Schneider, D., Schleichert, H., Strehl, U., Uhlmann, C., Blankenhorn, V., Fröscher, W., & Birbaumer, N. (1996). Self-regulation of slow cortical potentials in epilepsy: A retrial with analysis of influencing factors. Epilepsy Research, 25(3), 269-276. https://doi.org/10.1016/s0920-1211(96)00082-4
Kotchoubey, B., Schneider, D., Uhlmann, C., Schleichert, H., & Birbaumer, N. (1997). Beyond habituation: Long-term repetition effects on visual event-related potentials in epileptic patients. Electroencephalography and Clinical Neurophysiology, 103(4), 450-456. https://doi.org/10.1016/s0013-4694(97)00026-6
Kotchoubey, B., Strehl, U., Holzapfel, S., Blankenhorn, V., Fröscher, W., & Birbaumer, N. (1999). Negative potential shifts and the prediction of the outcome of neurofeedback therapy in epilepsy. Clinical Neurophysiology, 110(4), 683-686. https://doi.org/10.1016/s1388-2457(99)00005-x
Kotchoubey, B., Strehl, U., Holzapfel, S., Schneider, D., Blankenhorn, V., & Birbaumer, N. (1999). Control of cortical excitability in epilepsy. Advances in Neurology, 81, 281-290.
Kotchoubey, B., Strehl, U., Uhlmann, C., Holzapfel, S., Konig, M., Fröscher, W., Blankenhorn, V., & Birbaumer, N. (2001). Modification of slow cortical potentials in patients with refractory epilepsy: A controlled outcome study. Epilepsia, 42(3), 406-416.
Kropotov, J. D. (2009). Quantitative EEG, event-related potentials and neurotherapy. Academic Press.
Landisman, C. E., & Connors, B. W. (2005). Long-term modulation of electrical synapses in the mammalian thalamus. Science, 310(5755), 1809-1813. https://doi.org/10.1126/science.1114655
Lin, M. T., & Beal, M. F. (2006). Mitochondrial dysfunction and oxidative stress in neurodegenerative diseases. Nature, 443(7113), 787–795. https://doi.org/10.1038/nature05292
Lopes da Silva, F. (1991). Neural mechanisms underlying brain waves: from neural membranes to networks. Electroencephalography and Clinical Neurophysiology, 79(2), 81–93. https://doi.org/10.1016/0013-4694(91)90044-5
López-Doménech, G., Higgs, N. R., Vaccaro, V., Roš, H., Arancibia-Cárcamo, I. L., & Kittler, J. T. (2016). Loss of dendritic complexity precedes neurodegeneration in a mouse model with disrupted mitochondrial dynamics. Cell Reports, 17(2), 317–327. https://doi.org/10.1016/j.celrep.2016.08.073
Lopez-Otin, C., Blasco, M. A., Partridge, L., Serrano, M., & Kroemer, G. (2013). The hallmarks of aging. Cell, 153(6), 1194–1217. https://doi.org/10.1016/j.cell.2013.05.039
Meshorer et al. (2002). Alternative splicing and neuritic mRNA translocation under long-term neuronal hypersensitivity. Science, 295(5554), 508-512. https://doi.org/10.1126/science.1066752
Molenberghs, P., Cunnington, R., & Mattingley, J. B. (2011). Brain regions with mirror properties: A meta-analysis of 125 human fMRI studies. Neuroscience & Biobehavioral Reviews, 36(1), 341-349. https://doi.org/10.1016/j.neubiorev.2011.07.004
Munro, C. A., McCaul, M. E., Wong, D. F., Oswald, L. M., Zhou, Y., Brasic, J., Kuwabara, H., Kumar, A., Alexander, M., Ye, W., & Wand, G. S. (2006). Sex differences in striatal dopamine release in healthy adults. Biological Psychiatry, 59(10), 966-974. https://doi.org/10.1016/j.biopsych.2006.01.008
Mölle, M., Marshall, L., Gais, S., & Born, J. (2002). Grouping of spindle activity during slow oscillations in human non-rapid eye movement sleep. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 22(24), 10941–10947. https://doi.org/10.1523/JNEUROSCI.22-24-10941.2002
Nagano, S., & Araki, T. (2021). Axonal transport and local translation of mRNA in neurodegenerative diseases. Frontiers in Molecular Neuroscience, 14, 697973. https://doi.org/10.3389/fnmol.2021.697973
Nash, J. M. (2011). The gift of mimicry. Your brain: A user's guide. Time.
Niedermeyer, E. (1999). Historical aspects. In E. Niedermeyer & F. Lopes da Silva (Eds.), Electroencephalography: Basic principles, clinical applications, and related fields (4th ed.). Williams and Wilkins.
Niedermeyer, E., & Lopes da Silva, F. (Eds.). (1999). Electroencephalography: Basic principles, clinical applications, and related fields (4th ed.). Williams and Wilkins.
Othmer, S., & Othmer, S. (2020). Toward a theory of infra-low frequency neurofeedback. In H. W. Kirk (Ed.), Restoring the Brain. Routledge, eBook ISBN 9780429275760
Otto, D., & Reiter, L. (1984). Developmental changes in slow cortical potentials of young children with elevated body lead burden: Neurophysiological considerations. Annals of the New York Academy of Sciences, 425(1), 377-383. https://doi.org/10.1111/j.1749-6632.1984.tb23559.x
Pulvermüller, F., Mohr, B., Schleichert, H., & Veit, R. (2000). Operant conditioning of left-hemispheric slow cortical potentials and its effect on word processing. Biological Psychology, 53(2-3), 177-215. https://doi.org/10.1016/S0301-0511(00)00046-6
Radley, J. J., Arias, C. M., & Sawchenko, P. E. (2006). Regional differentiation of the medial prefrontal cortex in regulating adaptive responses to acute emotional stress. The Journal of Neuroscience, 26(50), 12967-12976. https://doi.org/10.1523/JNEUROSCI.4297-06.2006
Raichle, M. E., & Gusnard, D. A. (2002). Appraising the brain's energy budget. Proceedings of the National Academy of Sciences, 99(16), 10237-10299. https://doi.org/10.1073/pnas.172399499
Rapanelli, M., Frick, L. R., & Zanutto, B. S. (2011). Learning an operant conditioning task differentially induces gliogenesis in the medial prefrontal cortex and neurogenesis in the hippocampus. PLoS ONE, 6(2), e14713. https://doi.org/10.1371/journal.pone.0014713
Rockstroh, B., Elbert, T., Birbaumer, N., Wolf, P., Düchting-Röth, A., Reker, M., Daum, I., Lutzenberger, W., & Dichgans, J. (1993). Cortical self-regulation in patients with epilepsies. Epilepsy Research, 14(1), 63–72. https://doi.org/10.1016/0920-1211(93)90075-i
Ros, T., Munneke, M. A. M., Ruge, D., Gruzelier, J. H., & Rothwell, J. C. (2010). Endogenous control of waking brain rhythms induces neuroplasticity in humans. European Journal of Neuroscience, 31(4), 770–778. https://doi.org/10.1111/j.1460-9568.2010.07100.x
Sarnthein, J., Petsche, H., Rappelsberger, P., Shaw, G. L., & von Stein, A. (1998). Synchronization between prefrontal and posterior association cortex during human working memory. Proceedings of the National Academy of Sciences, 95(12), 7092-7096. https://doi.org/10.1073/pnas.95.12.7092
Schacter, D. L. (1977). EEG theta waves and psychological phenomena: A review and analysis. Biological Psychology, 5, 47-82. https://doi.org/10.1016/0301-0511(77)90028-x
Schneider, F., Rockstroh, B., Heimann, H., Lutzenberger, W., Mattes, R., Elbert, T., Birbaumer, N., & Bartels, M. (1992). Self-regulation of slow cortical potentials in psychiatric patients: Schizophrenia. Biofeedback and Self-Regulation, 17(4), 277-292. https://doi.org/10.1007/bf01000051
Schwartz, M. S., & Andrasik, F. (Eds.). (2003). Biofeedback: A practitioner's guide (3rd ed.). The Guilford Press.
Sebel, P. S., Lang, E., Rampil, I. J., White, P. F., Cork, R., Jopling, M., Smith, N. T., Glass, P. S., & Manberg, P. (1997). A multicenter study of bispectral electroencephalogram analysis monitoring anesthetic effect. Anesthesia and Analgesia, 84(4), 891-899. https://doi.org/10.1097/00000539-199704000-00035
Shibasaki, H., & Hallett, M. (2006). What is the Bereitschaftspotential? Clinical Neurophysiology, 117(11), 2341-2356. https://doi.org/10.1016/j.clinph.2006.04.025
Siniatchkin, M., Hierundar, A., Kropp, P., Kuhnert, R., Gerber, W-D., & Stephani, U. (2000). Self-regulation of slow cortical potentials in children with migraine: An exploratory study. Applied Psychophysiology & Biofeedback, 25(1), 13-32. https://doi.org/10.1023/a:1009581321624
Sitaram, R., Ros, T., Stoeckel, L., Haller, S., Scharnowski, F., Lewis-Peacock, J., Weiskopf, N., Blefari, M. L., Rana, M., Oblak, E., Birbaumer, N., & Sulzer, J. (2017). Closed-loop brain training: The science of neurofeedback. Nature Reviews Neuroscience, 18(2), 86–100. https://doi.org/10.1038/nrn.2016.164
Smith, M. L. (2013). Infra-slow fluctuation training; On the down-low in neuromodulation. NeuroConnections.
Speckmann, E.-J., & Elger, C. E. (1984). The neurophysiological basis of epileptic activity: A condensed overview. In R. Degen & E. Niedermeyer (Eds.), Epilepsy, sleep, and sleep deprivation (pp.23-34). Elsevier. PMID: 1760082
Speckmann, E.-J., & Elger, C. E. (1999). Introduction to the neurophysiological basis of the EEG and DC potentials. In E. Niedermeyer & F. Lopes da Silva (Eds.), Electroencephalography: Basic principles, clinical applications, and related fields (4th ed.). Williams and Wilkins.
Stahl, S. M. (2008). Stahl's essential psychopharmacology: Neuroscientific basis and practical applications (3rd ed.). Cambridge University Press.
Steriade, M. (2005). Cellular substrates of brain rhythms. In E. Niedermeyer, & F. Lopes da Silva (Eds.). Electroencephalography: Basic principles, clinical applications, and related fields (5th ed.). Lippincott Williams & Wilkins.
Steriade, M., Nuñez, A., & Amzica, F. (1993). Intracellular analysis of relations between the slow (< 1 Hz) neocortical oscillation and other sleep rhythms of the electroencephalogram. The Journal of Neuroscience: The Official Journal of the Society for Neuroscience, 13(8), 3266–3283. https://doi.org/10.1523/JNEUROSCI.13-08-03266.1993
Sterman, M. B. (1996). Physiological origins and functional correlates of EEG rhythmic activities: implications for self-regulation. Biofeedback and Self-regulation, 21(1), 3–33. https://doi.org/10.1007/BF02214147
Sterman, M. B. (2000). EEG markers for attention deficit disorder: Pharmacological and neurofeedback applications. Child Study Journal, 30(1), 1-24.
Stern, R. M., Ray, W. J., & Quigley, K. S. (2001). Psychophysiological recording (2nd ed.). Oxford University Press.
Strehl, U. (2009). Slow cortical potentials neurofeedback. Journal of Neurotherapy, 13(2), 117–126. https://doi.org/10.1080/10874200902885936
Strehl, U., Aggensteiner, P., Wachtlin, D., Brandeis, D., Albrecht, B., Arana, M., Bach, C., Banaschewski, T., Bogen, T., Flaig-Röhr, A., Freitag, C. M., Fuchsenberger, Y., Gest, S., Gevensleben, H., Herde, L., Hohmann, S., Legenbauer, T., Marx, A. M., Millenet, S., Pniewski, B., … Holtmann, M. (2017). Neurofeedback of slow cortical potentials in children with Attention-Deficit/Hyperactivity Disorder: A multicenter randomized trial controlling for unspecific effects. Frontiers in Human Neuroscience, 11, 135. https://doi.org/10.3389/fnhum.2017.00135
Streit, W. J. (2006). Microglial senescence: Does the brain's immune system have an expiration date? Trends in Neurosciences, 29(9), 506-510. https://doi.org/10.1016/j.tins.2006.07.001
Thompson, M., & Thompson, L. (2009). Asperger's syndrome intervention: Combining neurofeedback, biofeedback, and metacognition. In T. H. Budzynski, H. K. Budzynski, J. R. Evans, & A. Abarbanel (Eds.). Introduction to quantitative EEG and neurofeedback (2nd ed.). Academic Press.
Thompson, M., & Thompson, L. (2015). The neurofeedback book: An introduction to basic concepts in applied psychophysiology (2nd ed.). Association for Applied Psychophysiology and Biofeedback.
Vardalaki, D., Chung, K., & Harnett, M. T. (2022). Filopodia are a structural substrate for silent synapses in adult neocortex. Nature, 612(7939), 323-327. https://doi.org/10.1038/s41586-022-05483-6
Vernon, D., Egner, T., Cooper, N., Compton, T., Neilands, C., Sheri, A., & Gruzelier, J. (2003). The effect of training distinct neurofeedback protocols on aspects of cognitive performance. International Journal of Psychophysiology: Official Journal of the International Organization of Psychophysiology, 47(1), 75–85. https://doi.org/10.1016/s0167-8760(02)00091-0
von Bartheld, C. S., Bahney, J., & Herculano-Houzel, S. (2016). The search for true numbers of neurons and glial cells in the human brain: A review of 150 years of cell counting. The Journal of Comparative Neurology, 524(18), 3865-3895. https://doi.org/10.1002/cne.24040
von Stein, A., Rappelsberger, P., Sarnthein, J., & Petsche, H. (1999). Synchronization between temporal and parietal cortex during multimodal object processing in man. Cerebral Cortex, 9(2), 137-150. https://doi.org/10.1093/cercor/9.2.137
von Stein, A., & Sarnthein, J. (2000). Different frequencies for different scales of cortical integration: From local gamma to long range alpha/theta synchronization. International Journal of Psychophysiology, 38(3), 301-313. https://doi.org/10.1016/s0167-8760(00)00172-0
Voytek, B. (2013). Brain metrics: How measuring brain biology can explain the phenomena of mind. Scitable by Nature Education.
Warren, A. M., & McIlvane, W. J. (1998). Stimulus equivalence and the N400 effect. Poster presented at the 1998 Annual Meeting of the Cognitive Neuroscience Society in San Francisco, CA.
West, A. P., Shadel, G. S., & Ghosh, S. (2015). Mitochondria in innate immune responses. Nature Reviews Immunology, 15(11), 505–518. https://doi.org/10.1038/nri3850
Wilson, J. (2003). Biological foundations of human behavior. Wadsworth/Thompson Learning.
Wilson, V. E., Thompson, M., Thompson, L., Thompson, J., Fallahpour, K., & Linden, M. K. (2011). Introduction to biofeedback (Neurofeedback). In B. W. Strack, M. K. Linden, & V. S. Wilson (Eds.). Biofeedback & neurofeedback applications in sport psychology. Association for Applied Psychophysiology and Biofeedback.
Winn, P. (Ed.). (2001). Dictionary of biological psychology. Routledge.
Xu, T., Yu, X., Perlik, A., Tobin, W., Zweig, J., Tennant, K., Jones, T., & Zuo, Y. (2009). Rapid formation and selective stabilization of synapses for enduring motor memories. Nature, 462(7275), 915-919. https://doi.org/10.1038/nature08389
Yang, Y., Ge, W., Chen, Y., Zhang, Z., Shen, W., Wu, C., Poo, M., & Duan, S. (2003). Contribution of astrocytes to hippocampal long-term potentiation through release of D-serine. Proceedings of the National Academy of Sciences of the United States of America, 100(25), 15194-15199. https://doi.org/10.1073/pnas.2431073100
Zhang, H., Watrous, A., Patel, A., & Jacobs, J. (2018). Theta and alpha oscillations are traveling waves in the human neocortex. Neuron. https://doi.org/10.1016/j.neuron.2018.05.019