Have you ever noticed your palms getting sweaty before a job interview, even though the room was cool? This unit explores the fascinating world of electrodermal activity, where your skin reveals what your mind is processing. You will discover how the eccrine sweat glands on your palms and fingertips respond to emotional arousal, cognitive challenges, and even subtle shifts in attention.
Along the way, you will learn the difference between tonic and phasic electrodermal measures, understand why researchers prefer skin conductance over skin resistance, and explore the brain regions that orchestrate these responses. You will also discover why some people are "labiles" who show robust electrodermal responses while others are "stabiles" with more muted reactions, and what these individual differences mean for clinical practice.
This unit covers Three Electrodermal Measures, the Skin, Eccrine Sweat Glands, Brain Areas That Control Electrodermal Activity, Factors That Influence Electrodermal Activity, and Behavioral Correlates of Electrodermal Activity.
BCIA Blueprint Coverage: This unit addresses Descriptions of the most commonly employed biofeedback modalities: Electrodermal activity (III-A 2) and Structure and function of the autonomic nervous system (V-A 3).
The skin is the largest organ in the human body. Your skin mirrors attentional, defensive, and problem-solving processes through tonic and phasic electrodermal activity dependent on eccrine sweat glands. These exocrine glands produce about 600 mL of sweat per day to regulate body temperature via evaporation (Tortora & Derrickson, 2021).
Cross-section of human skin showing the epidermis, dermis, and hypodermis with eccrine sweat glands. Jiri Vratislavsky/Shutterstock.com.
"Social and behavioral scientists have found that tonic EDA is useful to investigate general states of arousal and/or alertness, and that the phasic SCR is useful to study multifaceted attentional processes, as well as individual differences in both the normal and abnormal spectrum" (Dawson et al., 2016, p. 237).
The palmar surface contains approximately 3,000 sweat glands per square inch, making it an ideal site for electrodermal monitoring.
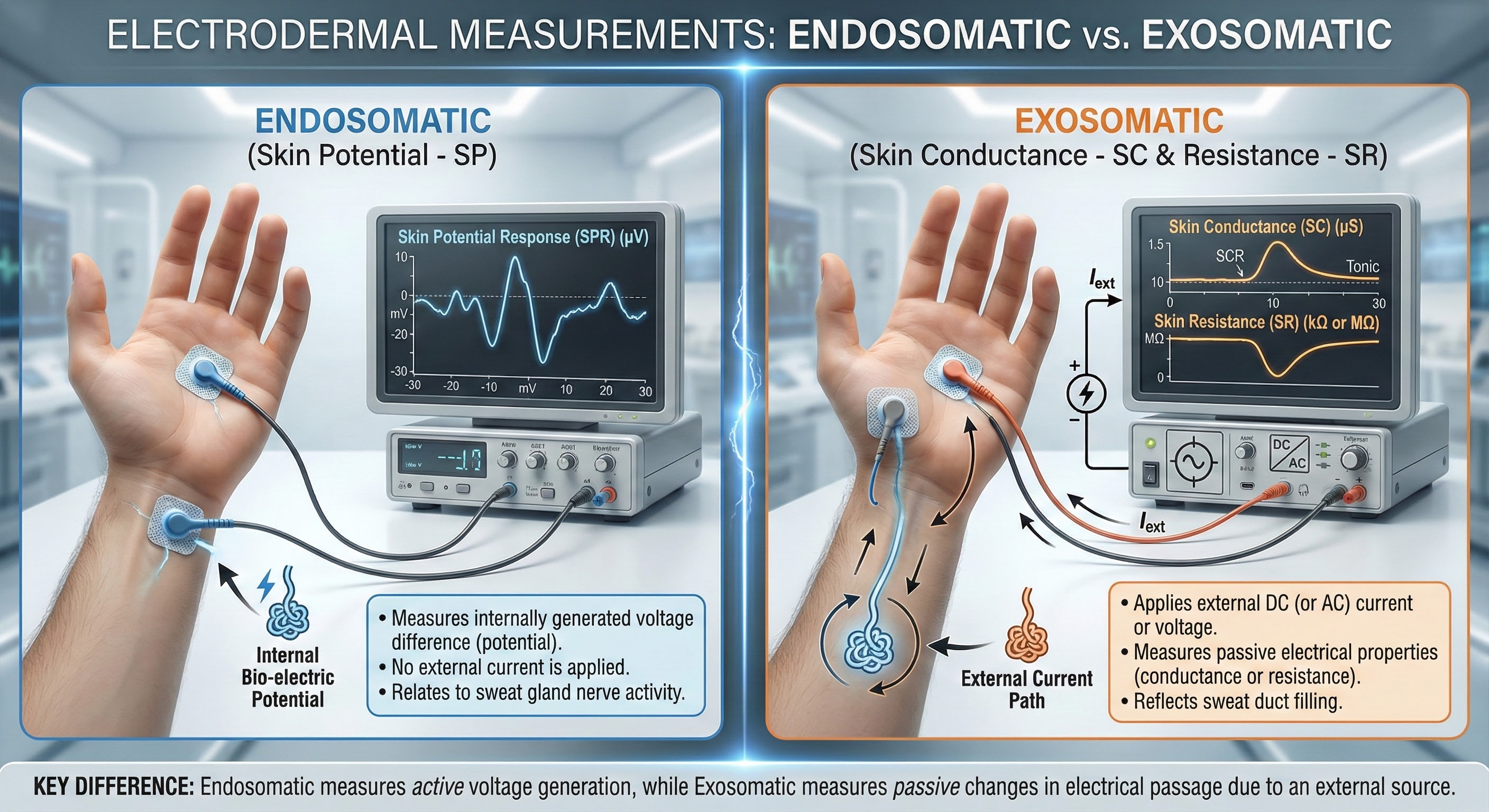
Practitioners measure electrodermal activity (EDA) using three methods: conductance, resistance, and potential. Conductance and resistance are measured exosomatically (from outside the body) by passing an electric current through the skin. Skin potential is measured endosomatically (from inside the body).
Active vs. Passive Measurement: This infographic illustrates the two primary methods of electrodermal recording. The left panel depicts Endosomatic measurement (Skin Potential), where electrodes detect voltage generated naturally by the body. The right panel shows Exosomatic measurement (Skin Conductance/Resistance), which requires an external current source to measure the skin's passive electrical properties.
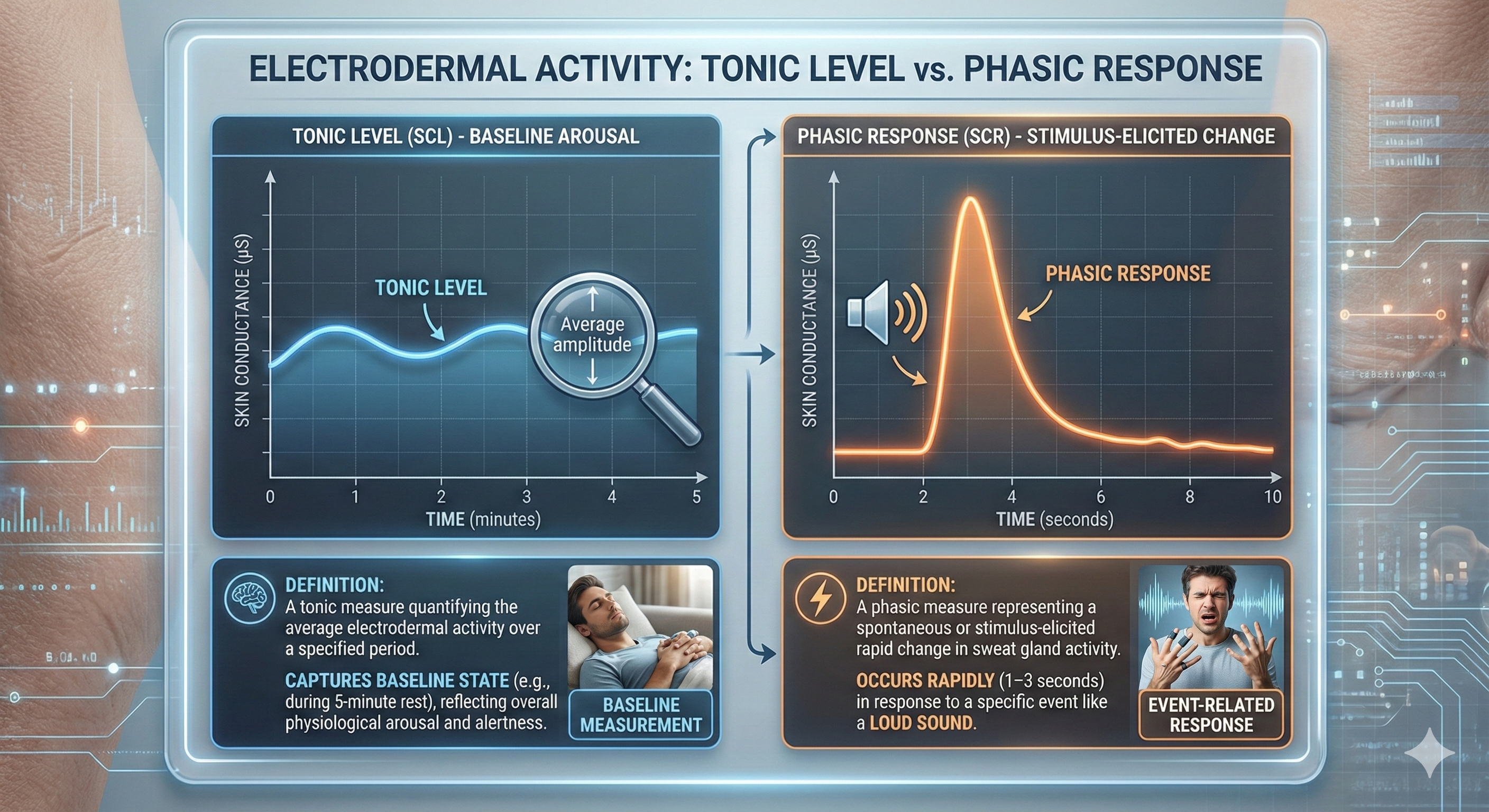
Before diving into these measures, you need to understand two fundamental concepts that apply to all electrodermal monitoring. Level is a tonic measure of electrodermal activity that quantifies the average amplitude over a specified period. Think of tonic measures as capturing your client's baseline state, like taking a snapshot of their overall arousal level during a 5-minute rest period.
Response is a phasic measure of EDA representing a spontaneous or stimulus-elicited change in sweat gland activity. A stimulus like a loud sound can produce an electrodermal response in 1-3 seconds (Andreassi, 2007). This is blazingly fast compared to the 20-30-second response time for skin temperature, making EDA particularly useful when you want to track moment-to-moment changes in arousal.
The Two Sides of EDA: This visual guide contrasts Tonic Level (your steady baseline arousal) with Phasic Response (immediate reactions to stimuli), illustrating how electrodermal activity measures both background physiological states and rapid spikes in sweat gland activity.
Skin Conductance: The Preferred Measure
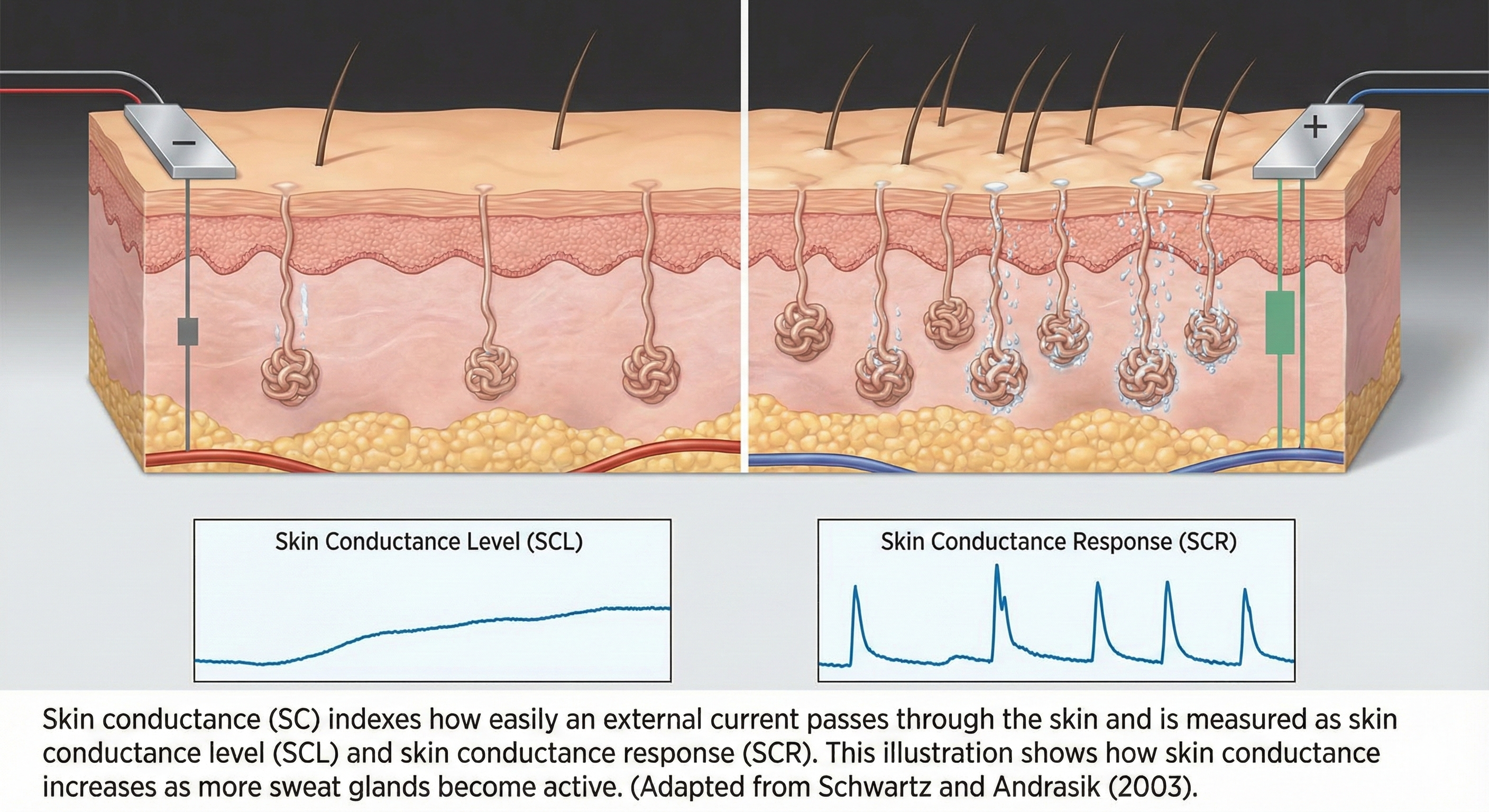
Skin conductance (SC) indexes how easily an external current passes through the skin and is measured as skin conductance level (SCL) and skin conductance response (SCR). The illustration below, adapted from Schwartz and Andrasik (2003), shows how skin conductance increases as more sweat glands become active.
This illustration demonstrates how increased sweat gland activity facilitates electrical current flow through the skin. The bottom panels distinguish between tonic Skin Conductance Level (SCL) and phasic Skin Conductance Response (SCR) waveforms.
Below is a BioGraph Infiniti display of resting skin conductance level.
Skin Resistance: The Historical Measure
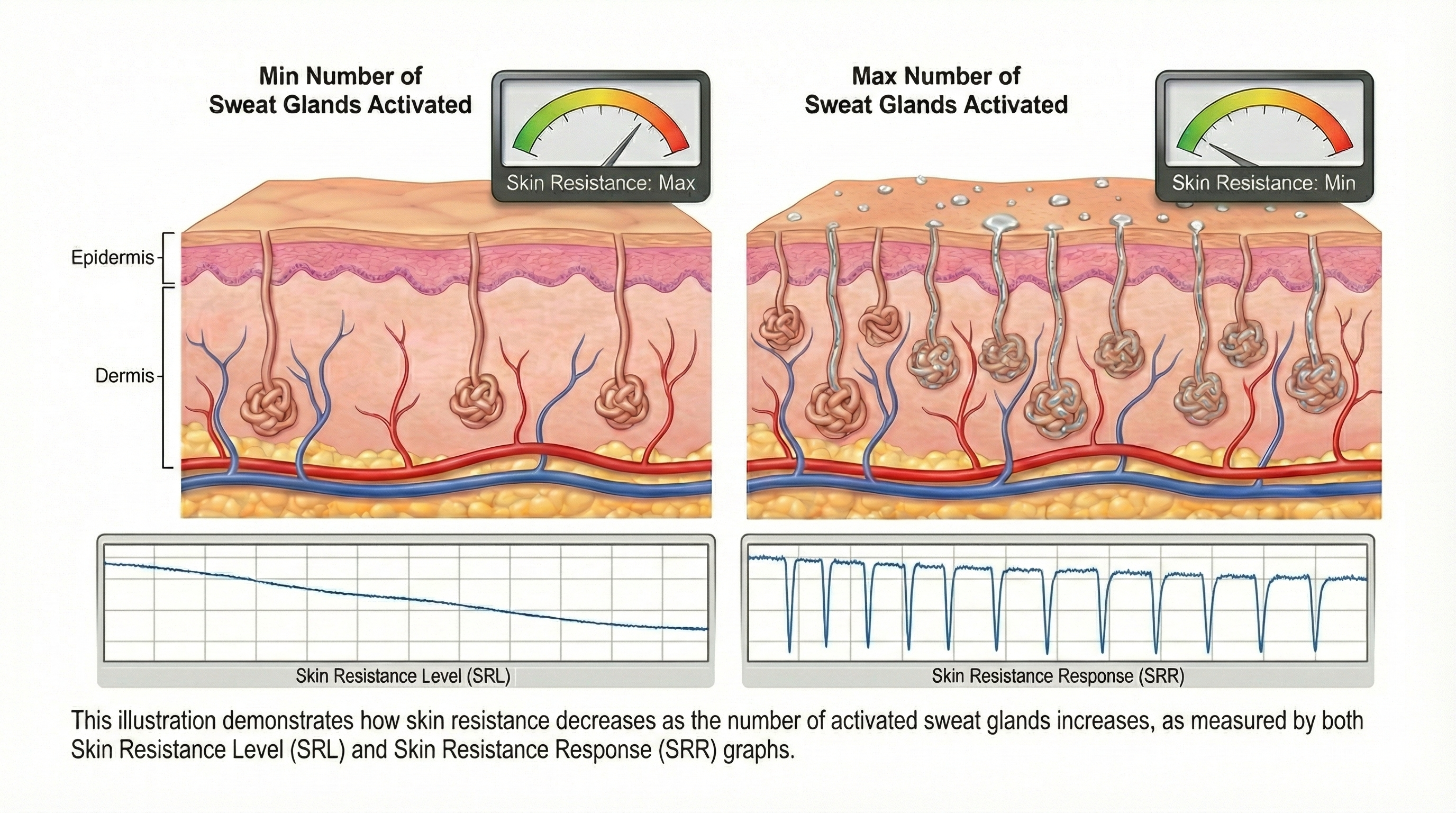
Skin resistance (SR), also called galvanic skin response (GSR), reflects opposition to external current movement and is measured as skin resistance level (SRL) and skin resistance response (SRR). The illustration below, adapted from Schwartz and Andrasik (2003), shows how skin resistance decreases as more sweat glands activate.
This diagram illustrates the inverse relationship between perspiration and electrical resistance: as sweat gland activation increases (right), skin resistance decreases, resulting in measurable spikes in conductivity known as the Skin Resistance Response (SRR).
Skin conductance is the reciprocal of skin resistance (SC = 1/SR). Researchers prefer skin conductance because it is more normally distributed than skin resistance and increases linearly as the sympathetic nervous system activates more sweat glands (Andreassi, 2007). This linear relationship makes skin conductance easier to interpret in clinical and research settings.
Skin Potential: The Endosomatic Measure
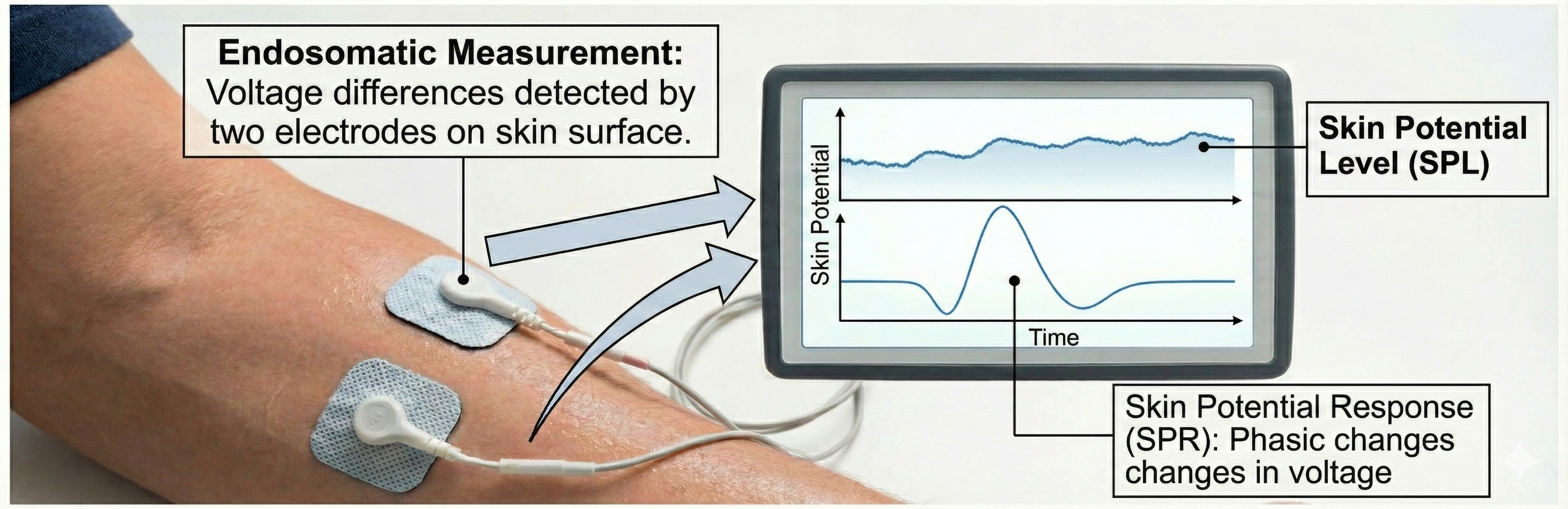
Skin potential is monitored endosomatically (from within the body) by detecting voltage differences between two electrodes on the skin surface. Skin potential is measured as skin potential level (SPL) and skin potential response (SPR). The illustration below, adapted from Schwartz and Andrasik (2003), depicts the relationship between skin potential and sweat gland activity.
An infographic illustrating how voltage differences between skin surface electrodes are monitored to track baseline Skin Potential Level (SPL) and the phasic, triphasic Skin Potential Response (SPR).
Three Electrodermal Measures
EDA can be measured three ways: skin conductance (how easily current flows), skin resistance (opposition to current flow), and skin potential (voltage differences). Each measure has tonic (level) and phasic (response) components. Researchers prefer skin conductance because it increases linearly with sympathetic activation and has better statistical properties than skin resistance.
Comprehension Questions: Electrodermal Measures
Why do researchers prefer skin conductance over skin resistance for measuring electrodermal activity?
What is the fundamental difference between exosomatic and endosomatic measurement methods?
How would you explain the difference between tonic and phasic measures to a client new to biofeedback?
Why is electrodermal activity considered a faster indicator of arousal changes than skin temperature?
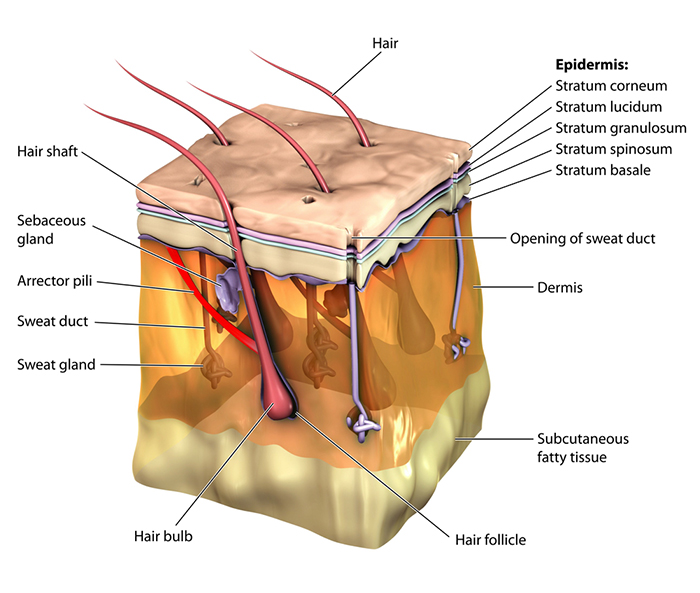
The Skin: Your Body's Largest Organ
The protective skin boundary consists of three main layers: the epidermis (outer layer), dermis (inner layer), and hypodermis. Understanding these layers helps you appreciate where electrodermal activity originates.
The epidermis comprises five layers: stratum corneum, stratum lucidum, stratum granulosum, stratum spinosum, and stratum germinativum. The dermis contains blood and lymph vessels, smooth muscle, and sebaceous and sweat glands. The secretory portion of sweat glands is located in the hypodermis, a region consisting of connective tissue below the dermis that contains blood and lymph vessels.
The three layers of human skin. The epidermis provides protection, the dermis contains blood vessels and glands, and the hypodermis houses the secretory portions of sweat glands. Blamb/Shutterstock.com.
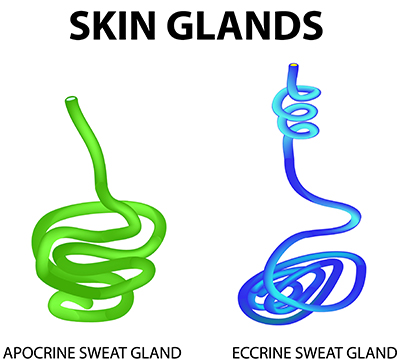
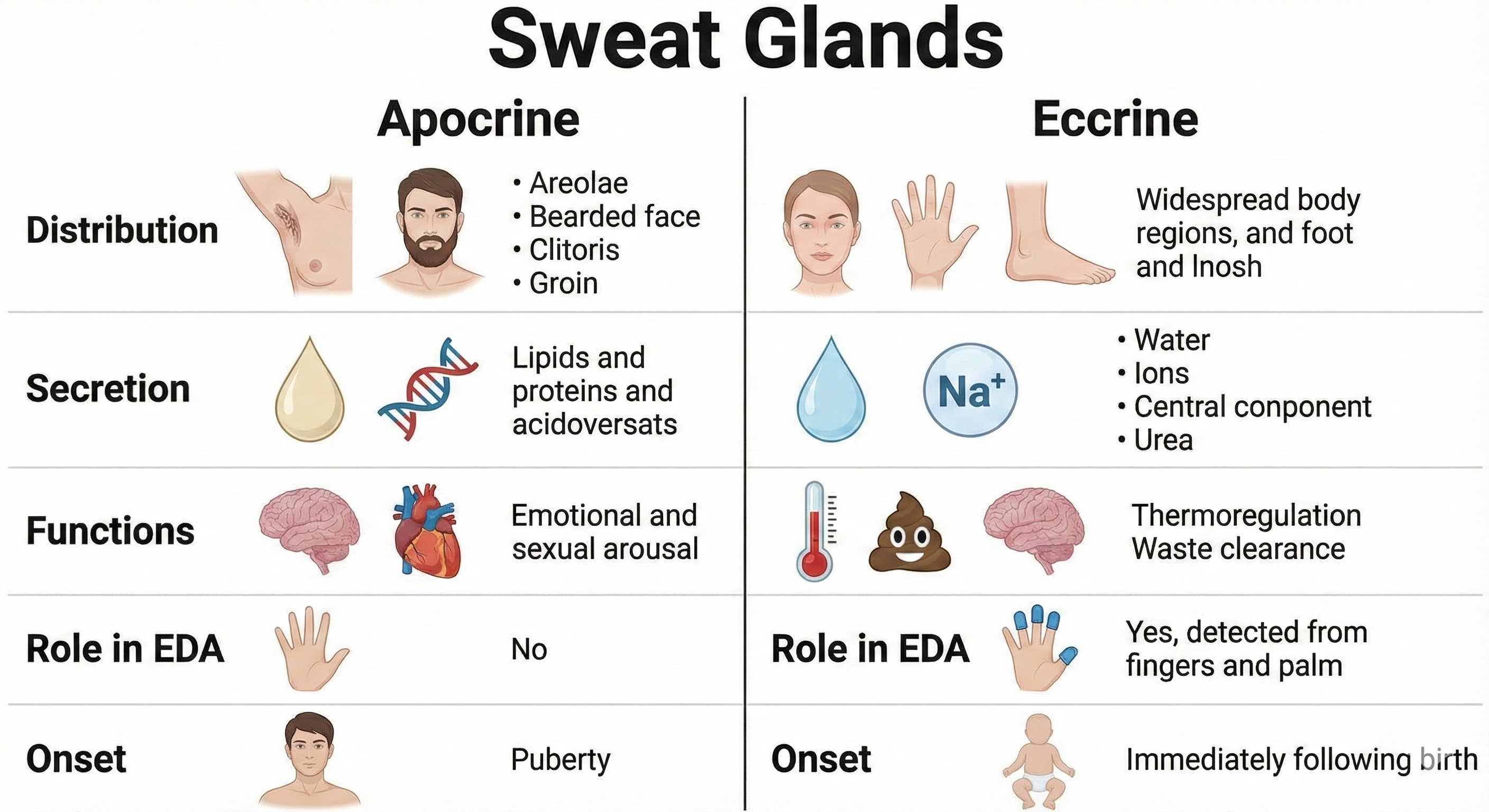
The human skin contains three-to-four million sudoriferous glands (sweat glands), including apocrine and eccrine sweat glands (Tortora & Derrickson, 2021). These two types of sweat glands serve very different functions.
Apocrine Sweat Glands: Not Involved in EDA
Apocrine sweat glands have larger-diameter ducts and lumens than eccrine sweat glands (Tortora & Derrickson, 2021). They usually open into hair follicles and are mainly distributed in the armpits, bearded male facial areas, breast areolae, and genital region. They secrete sweat using exocytosis. White- or yellow-colored apocrine sweat produces a musky odor when combined with skin surface bacteria. Apocrine sweat begins during puberty and is released during emotional reactions and sexual activity. However, apocrine sweat glands play no role in thermoregulation or the electrodermal activity you measure in biofeedback.
Comparison of apocrine and eccrine sweat glands. Apocrine glands are associated with hair follicles and produce scented sweat, while eccrine glands open directly onto the skin surface. Timonina/Shutterstock.com.
Eccrine Sweat Glands: The Source of EDA
Eccrine sweat glands are mainly responsible for EDA and are distributed across the body surface, especially on the forehead, palms, and soles. They are more numerous than apocrine sweat glands (Tortora & Derrickson, 2021). Eccrine sweat glands are not found in the clitoris, labia minora, glans penis, eardrums, lip margins, nail beds of the fingers and toes, or the outer ear. A square inch of the palmar surface may contain about 3,000 sweat glands (Jacob & Francone, 1970). Cadaver studies indicate that the entire body may have 2-5 million sweat glands (Fowles, 1986).
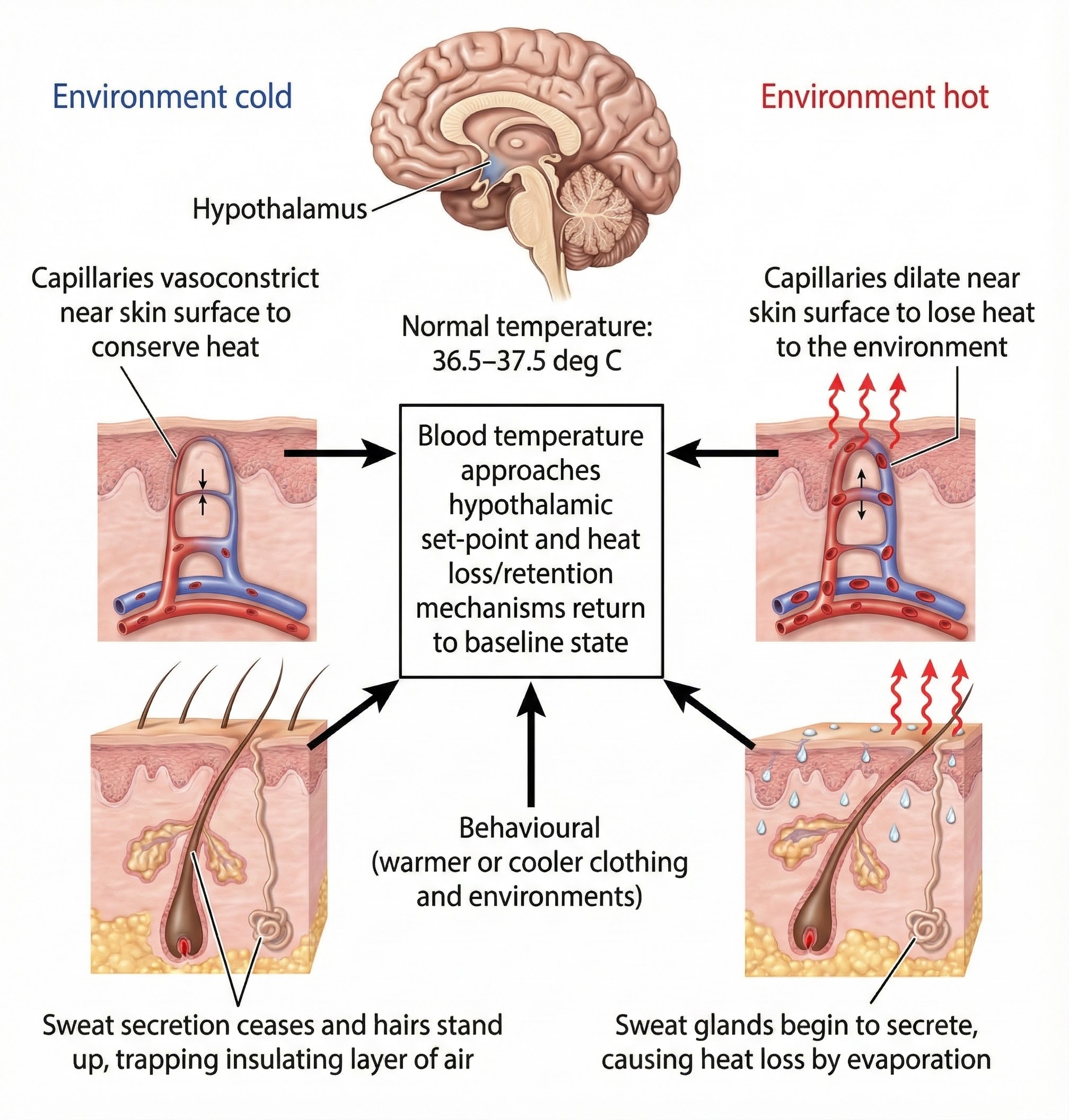
Thermoregulation: Keeping Your Cool
Peripheral and central thermoreceptors monitor body temperature. They report temperature changes to the preoptic area of the hypothalamus and other brain regions (Breedlove & Watson, 2020). When body temperature rises, the preoptic area activates the heat-loss center and suppresses the heat-promoting center. The heat-loss center signals skin blood vessels to dilate to radiate heat into the environment. The hypothalamus activates sympathetic nerves that innervate eccrine sweat glands to initiate thermoregulation (temperature control) through evaporative cooling. This mechanism is active after birth (Tortora & Derrickson, 2021).
The hypothalamus coordinates thermoregulation by monitoring body temperature and activating appropriate cooling or heating mechanisms. Adapted from Blamb/Shutterstock.com.
Thermoregulatory sweating starts at the forehead and scalp, and last appears on the palms and soles. This order is not accidental since forehead and scalp sweating sheds the largest amount of heat from the body. In contrast, emotional sweating in response to fear or anxiety begins at the palms, soles, and armpits. Apocrine sweat glands also participate in emotional sweating.
Emotional Sweating: Your Palms Tell the Story
While all eccrine sweat glands respond to cognitive activity, emotion, and temperature, palmar and plantar sweat glands appear more responsive to emotional stimuli because of their higher density (about 1,000 glands per cm² compared with 100-200 per cm² on the trunk and limbs). Palmar and plantar sweating seems specialized for grasping objects, increasing tactile sensitivity, and protecting skin from damage (Dawson et al., 2016; Hugdahl, 1995). These changes support the human fight-or-flight response.
Regional differences in sweat gland density and function explain why the palms and soles are preferred sites for electrodermal monitoring.
Why Palmar Sites Are Preferred
Consider why you place electrodermal sensors on your client's fingers or palms rather than their forearm. The palms have approximately 1,000 sweat glands per square centimeter, compared to only 100-200 per square centimeter on the trunk. This higher density means you get a stronger, more reliable signal. Additionally, palmar sweating responds primarily to emotional and cognitive stimuli rather than temperature, making it ideal for tracking your client's psychological responses during biofeedback training.
The Skin and Sweat Glands
The skin has three layers: epidermis, dermis, and hypodermis. Eccrine sweat glands, responsible for EDA, are densest on the palms, soles, and forehead. Thermoregulatory sweating begins on the head, while emotional sweating begins on the palms and soles. The high density of palmar sweat glands (about 1,000 per cm²) makes the hands ideal for electrodermal monitoring because they respond robustly to emotional and cognitive stimuli.
Comprehension Questions: Skin and Sweat Glands
Why do thermoregulatory sweating and emotional sweating begin at different body locations?
How does the distribution of eccrine sweat glands explain why practitioners monitor EDA from the hands?
What role do apocrine sweat glands play in electrodermal activity measured during biofeedback?
Your client asks why you place sensors on their fingers instead of their forearm. How would you explain this choice?
Eccrine Sweat Gland Anatomy and Function
Structural Components
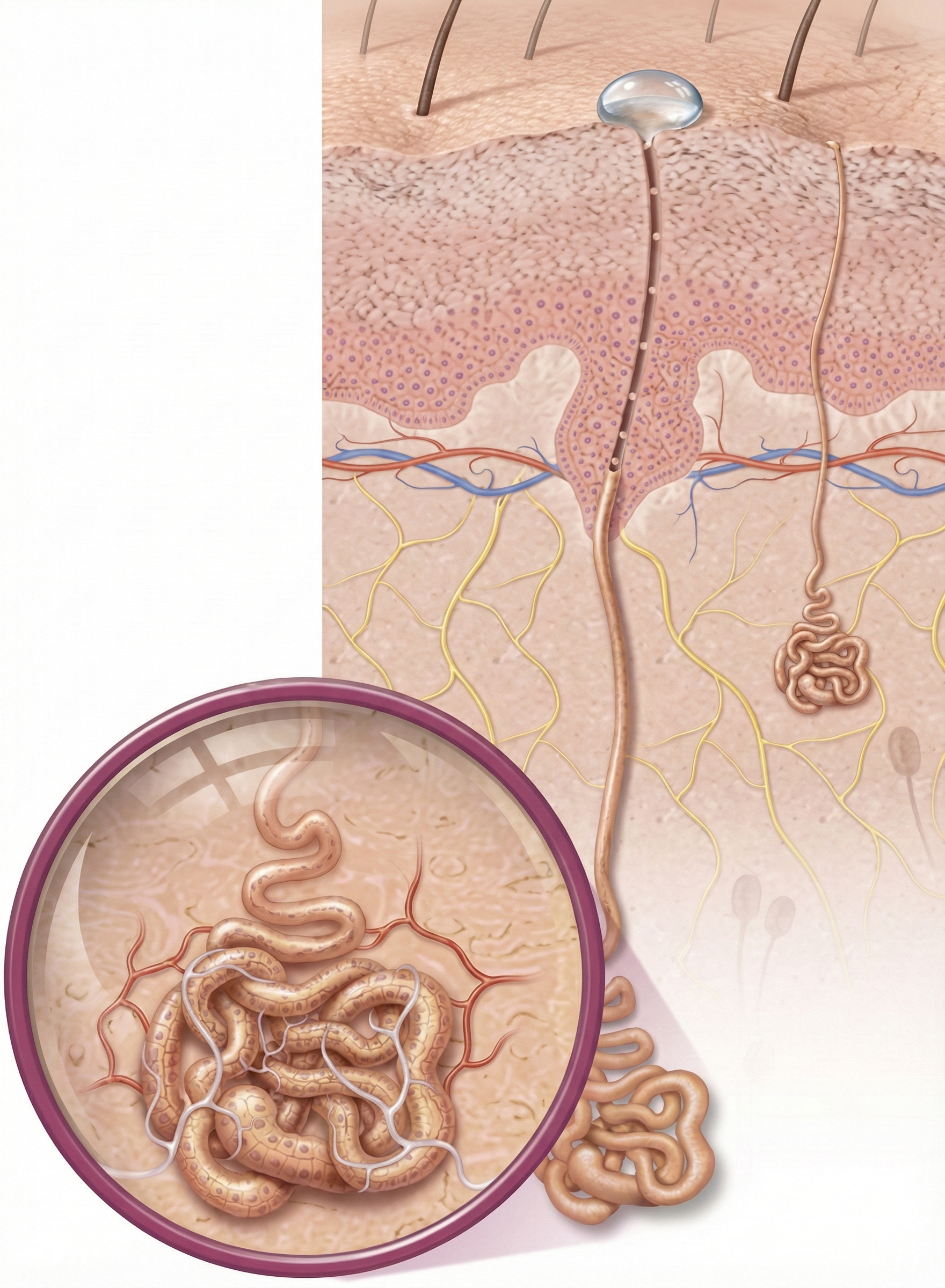
An eccrine sweat gland consists of a secretory portion and sweat duct. The secretory portion that produces sweat consists of coils arranged in a ball 0.3-0.4 mm in diameter. In the graphic below, sweat glands are shown in purple.
Structure of an eccrine sweat gland showing the coiled secretory portion and the duct leading to the skin surface. Tefi/Shutterstock.com.
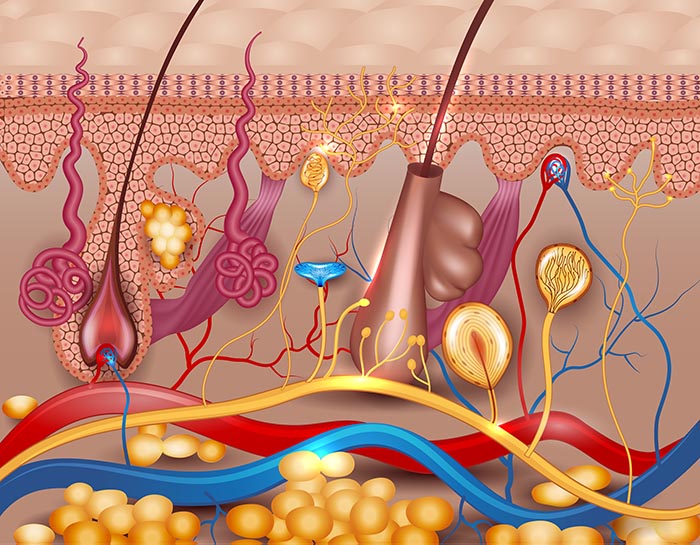
Sympathetic cholinergic fibers mainly innervate eccrine sweat glands. Unmyelinated cholinergic fibers are densest around the secretory portion. A few lie close to the duct. Researchers have also found nearby adrenergic fibers. Neurotransmitters like VIP may complement ACh and NE (Bach, 2014; Shields et al., 1987).
An infographic summarizing the physiological mechanisms behind Electrodermal Activity (EDA), highlighting the role of eccrine sweat glands, neurotransmitters, and signal characteristics.
Sweat gland coils are lined by myoepithelial cells that resemble smooth muscle cells. Myoepithelial cells may help produce spontaneous EDA. They are influenced by the hormones norepinephrine (NE) and possibly epinephrine (E). The sweat duct is a long tube that excretes sweat through a pore at the epidermis. Check out the Encyclopedia Britannica YouTube video Eccrine Glands.
The magnified area shows a sweat gland's secretory portion where sweat production begins.
EDA is typically recorded from the fingers and palmar surface of the hands. Different body areas (e.g., armpit) may respond differently to diverse stimuli. Only a few studies have monitored EDA from other regions, such as the forehead when training patients to inhibit motion sickness (Shaffer, Combatalade, Peper, & Meehan, 2016).
Sympathetic Control of Sweating
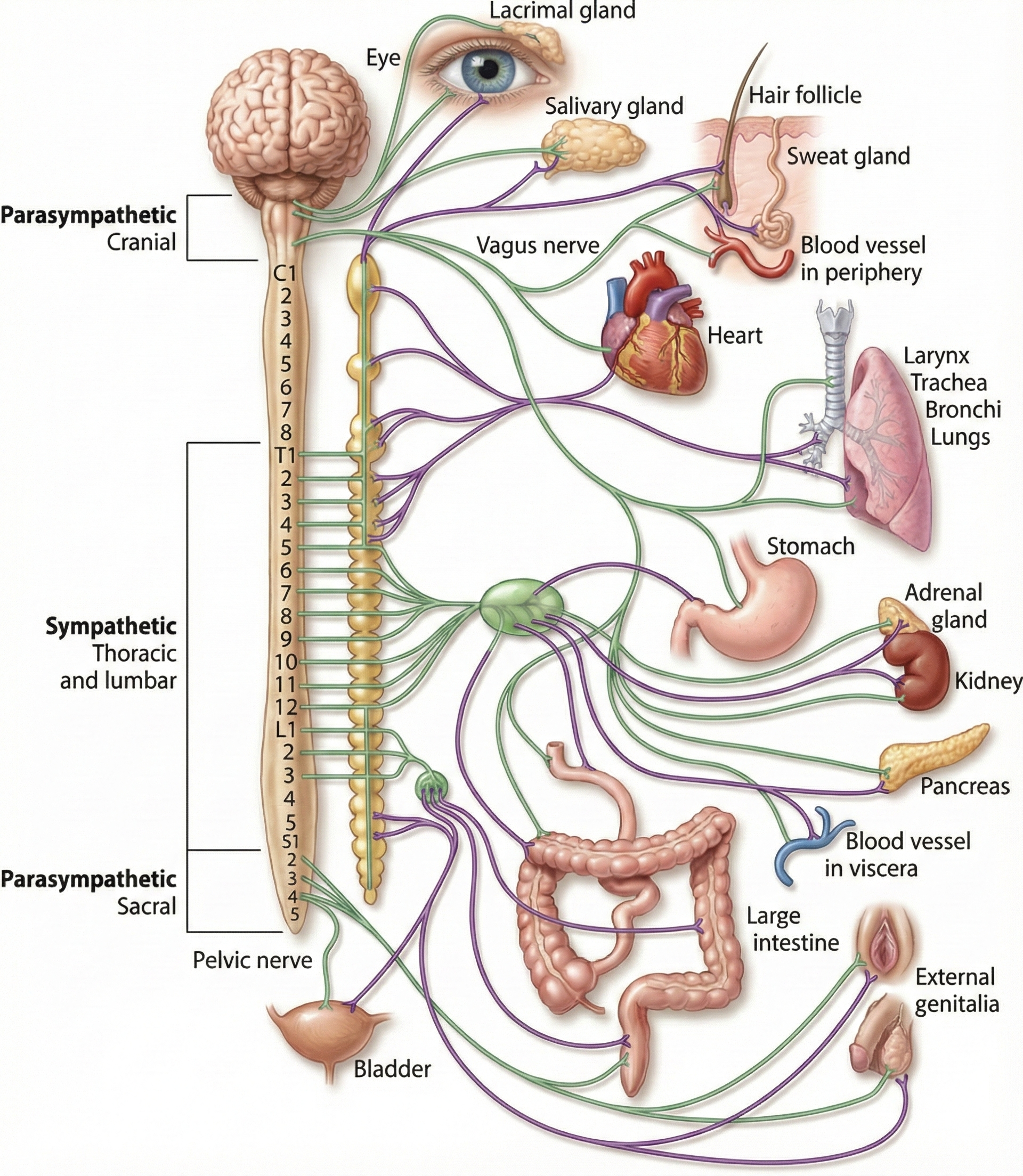
The sympathetic nervous system (SNS), the autonomic nervous system branch regulating activities that expend stored energy, primarily controls EDA. This view is supported by the strong correlation between sympathetic action potentials and skin conductance responses (SCRs) at average room temperature (Wallin, 1981). Increased SNS activation results in greater sweating from the palms.
EDA amplitude does not increase equally in both hands. Discrepancies between left- and right-hand values mainly reflect left/right brain function but may also signal pathology (Banks et al., 2012). Skin responses may also be affected by the particular dermatomes (areas of skin innervated by a single spinal nerve) activated.
A diagram illustrating the neural pathways of the sympathetic and parasympathetic divisions and their connections to major organs. Adapted from Josephine Wilson (2003).
Sweat Production
Human sweat begins as plasma-like precursor sweat. Sweat mainly comprises water, ions (Na+ and Cl-), amino acids, ammonia, glucose, lactic acid, urea, and uric acid (Tortora & Derrickson, 2021). The sweat duct reabsorbs sodium chloride from sweat.
Neurotransmitter Control of Eccrine Sweat Glands
When you feel your palms get sweaty before giving a presentation, a fascinating chain of chemical signals is at work beneath your skin. At the heart of this process is a single molecule that serves as the primary trigger for sweat production in humans: acetylcholine, a neurotransmitter that nerve cells release to communicate with sweat glands and other target tissues.
Here is where the story gets interesting. The eccrine sweat glands receive their commands from the sympathetic nervous system, the branch of your autonomic nervous system responsible for mobilizing your body during stressful or demanding situations. Throughout most of this system, nerve endings release a different chemical messenger called norepinephrine, a neurotransmitter typically associated with arousal and alertness. However, the nerves controlling eccrine sweat glands break this pattern entirely. Instead of releasing norepinephrine like their sympathetic neighbors, these nerve endings release acetylcholine (Hu, Converse, Lyons, & Hsu, 2018; Cui & Schlessinger, 2015). This makes eccrine sweat glands a notable exception to the usual rules of sympathetic nervous system signaling.
Once released, acetylcholine does not simply float around hoping to find a target. It binds to specific protein structures on the surface of secretory cells within the sweat gland. These structures are called muscarinic M3 receptors, specialized proteins embedded in cell membranes that recognize and respond specifically to acetylcholine. When acetylcholine locks onto these receptors, it sets off a cascade of events inside the cell. Calcium levels rise, which in turn activates ion channels (protein tunnels that allow charged particles to move across cell membranes) and transporters (proteins that actively shuttle substances from one side of a membrane to the other). These molecular machines work together to pump salt into the hollow center of the gland, known as the lumen. Water naturally follows the salt through a process called osmosis, and the result is sweat (Cui & Schlessinger, 2015; Klaka et al., 2017).
Scientists have confirmed the essential role of acetylcholine through elegant experiments. When researchers administer atropine, a drug that blocks muscarinic receptors and prevents acetylcholine from binding, sweating nearly stops completely. This holds true whether the sweating was triggered by heat or by psychological stress, demonstrating that acetylcholine is indispensable for normal eccrine sweat production (Machado-Moreira et al., 2012).
Supporting Players in Sweat Regulation
While acetylcholine runs the show, other chemical messengers contribute supporting roles. Eccrine sweat glands possess adrenergic receptors, cell surface proteins that respond to catecholamines such as adrenaline and noradrenaline (the hormonal forms of epinephrine and norepinephrine). These stress hormones can influence sweating, particularly during intense emotional experiences. However, they appear to fine-tune the system rather than replace the primary cholinergic drive (Hu et al., 2018; Harker, 2013). Think of catecholamines as volume knobs that can turn sweating up or down, while acetylcholine remains the power switch.
The regulatory picture grows even more complex when we consider co-transmitters, signaling molecules released alongside the primary neurotransmitter to modify or enhance its effects. Two peptides in particular, vasoactive intestinal peptide (VIP) and peptide histidine isoleucine, have been found in the nerve fibers surrounding eccrine glands. These peptides bind to secretory cells and have even been detected in sweat itself, suggesting they help regulate gland activity and local blood flow (Eedy, Johnston, Shaw, & Buchanan, 1990; Freeman, Waldorf, & Dover, 1992).
Additional local mediators likely add yet another layer of control. Nitric oxide, a small gaseous signaling molecule produced within tissues, along with various neuropeptides (small protein-like molecules used by neurons to communicate), probably contribute to the intrinsic regulation happening right at the gland itself. Despite this complexity, the central message remains clear: acetylcholine released from sympathetic cholinergic nerves is the main trigger for eccrine sweating in humans, with adrenergic and peptidergic signals providing additional modulation (Hu et al., 2018; Zancanaro, Merigo, Crescimanno, Orlandini, & Osculati, 1999).
The Electric Skin: Decoding the Signals Beneath the Surface
Imagine the sensation of your palms becoming clammy just before you have to speak in public, or the subtle jolt you feel when a sudden, loud noise startles you. These physical reactions are not merely symptoms of stress; they are readable data streams broadcast by your nervous system. This phenomenon is known as electrodermal activity (EDA), an umbrella term for all electrical properties of the skin. To understand this complex signal, researchers rely on two complementary frameworks: the poral valve model, which explains the mechanics at the skin level, and the poly-hierarchical neural model, which explains the control systems in the brain. We will first look at the machinery of the skin before exploring the neural "software" that drives it (Boucsein, 2012).
The Plumbing of Perception: Mechanics at the Skin Level
To understand how the skin generates an electrical signal, one must look at the body’s "hardware": the eccrine sweat glands. Concentrated densely on the palms and soles, these glands are the primary engine of EDA. According to the poral valve model, the skin’s outer layer, the stratum corneum, normally acts as a resistor—a dry shield that resists the flow of electricity. However, when the sympathetic nervous system activates, it triggers the secretory coil deep in the dermis to produce sweat. As this salty, conductive fluid rises up the sweat duct, it creates a path of least resistance through the skin’s barrier (Dawson et al., 2017).
This hydraulic action creates the most common measure of EDA, known as skin conductance. You can visualize the sweat ducts as variable resistors wired in parallel; the more ducts that fill with sweat, the higher the conductance. This relationship is linear, making skin conductance a highly reliable metric for gauging arousal (Fowles et al., 1981). However, the system is more complex than a simple pipe. The model proposes a functional poral valve at the skin's surface. When this valve opens to release sweat, conductance spikes. When the valve closes and sweat is reabsorbed, conductance recovers to baseline (Edelberg, 1993).
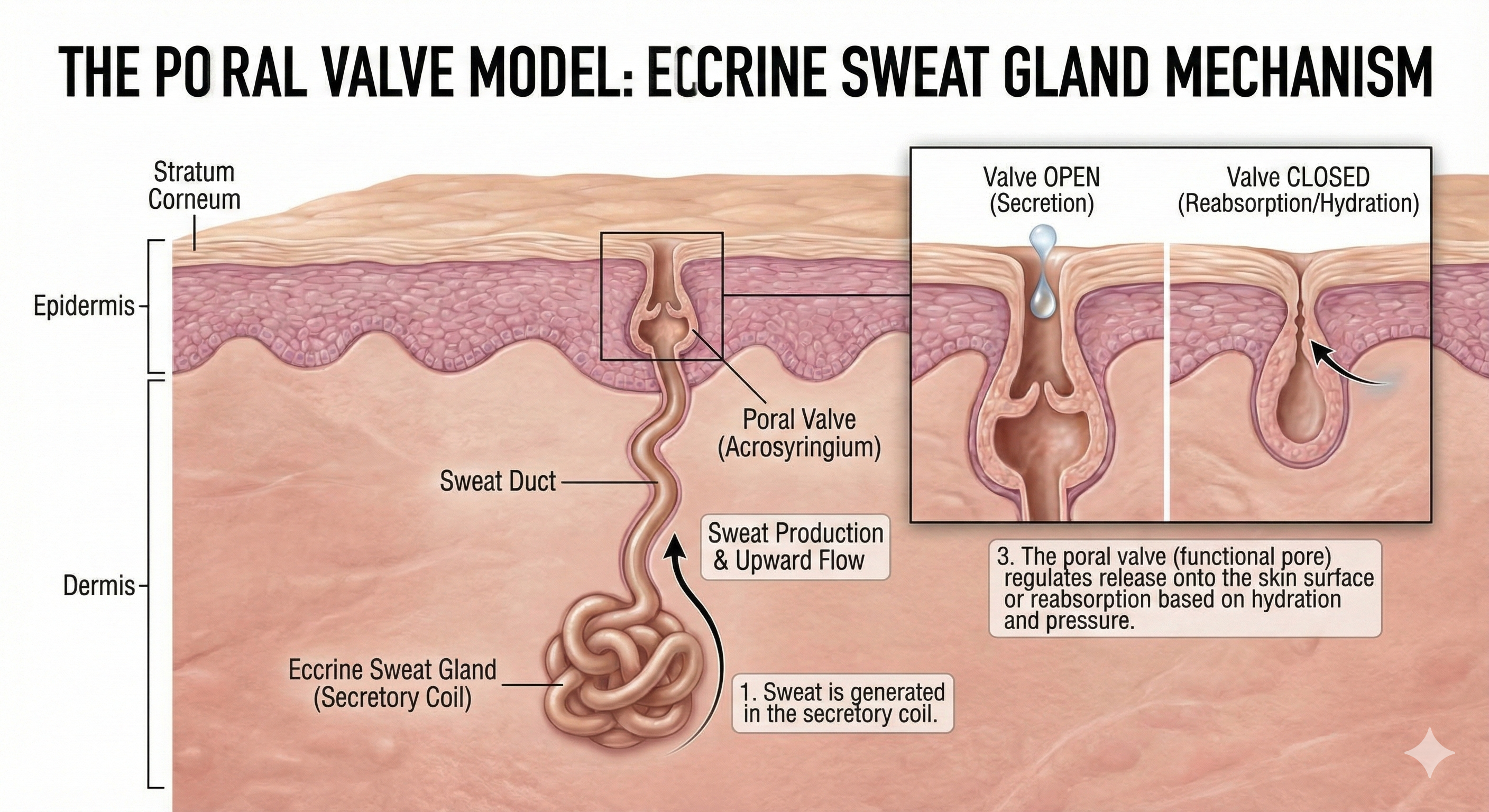
This graphic illustrates the Poral Valve Model, detailing the anatomical mechanism by which eccrine sweat glands regulate sweat production, secretion, and reabsorption in human skin.
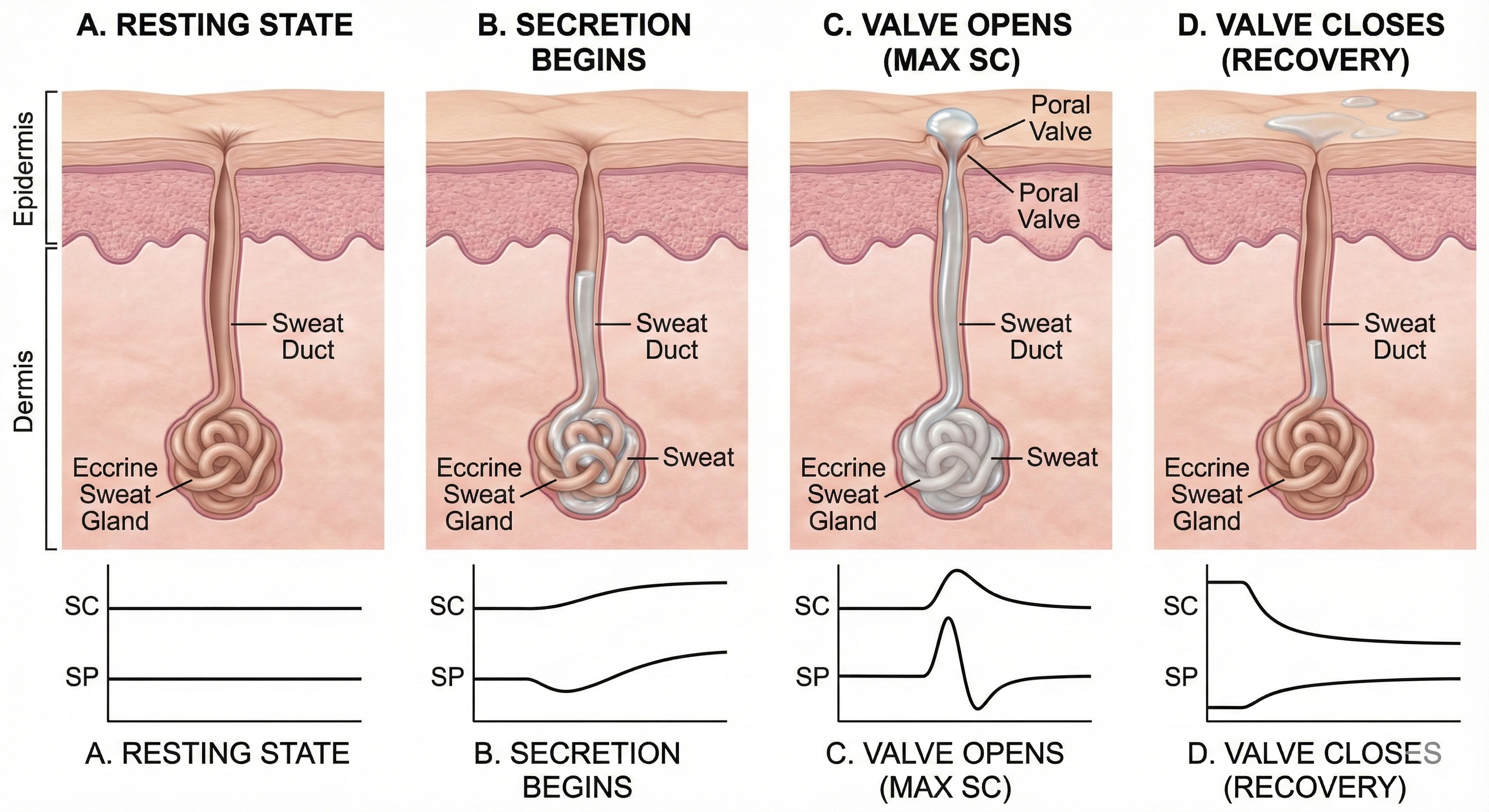
Edelberg (1993) explained the rapid shift from conductance to resistance in the sweat circuit model with his poral valve model. Initially, the sweat duct is empty, and the pore at the skin surface is closed (A). Sweat secretion rises in the duct, increasing skin conductance, and the pore remains closed (B). Sweat fills the duct until pressure forces the poral valve open, producing maximum skin conductance (C). Loss of sweat to the skin surface reduces the intraductal pressure needed to keep the poral valve open, causing it to close and conductance to decline rapidly (D).
The Poral Valve Model of Electrodermal Activity: This sequence illustrates the mechanical changes in an eccrine sweat gland that drive skin conductance (SC). (A) Baseline: The sweat duct is empty, and the surface pore is closed, resulting in low conductance. (B) Rising Conductance: Sweat secretion fills the duct, creating a conductive path, though the pore remains closed. (C) Peak Conductance: Pressure from the rising sweat forces the poral valve open, maximizing conductance. (D) Recovery: Sweat release relieves pressure, allowing the valve to close and conductance to rapidly decline.
While conductance measures how easily current flows, the skin also acts as a battery, generating its own voltage. This is known as skin potential. The model explains that skin potential is a biphasic signal—meaning it has both negative and positive components—driven by two different biological generators. The initial surge of sweat in the secretory coil creates a negative potential, while the epidermal barrier layer generates a positive potential (Boucsein, 2012). This duality allows researchers to distinguish between simple arousal and more complex defensive reflexes, something simple conductance cannot always reveal.
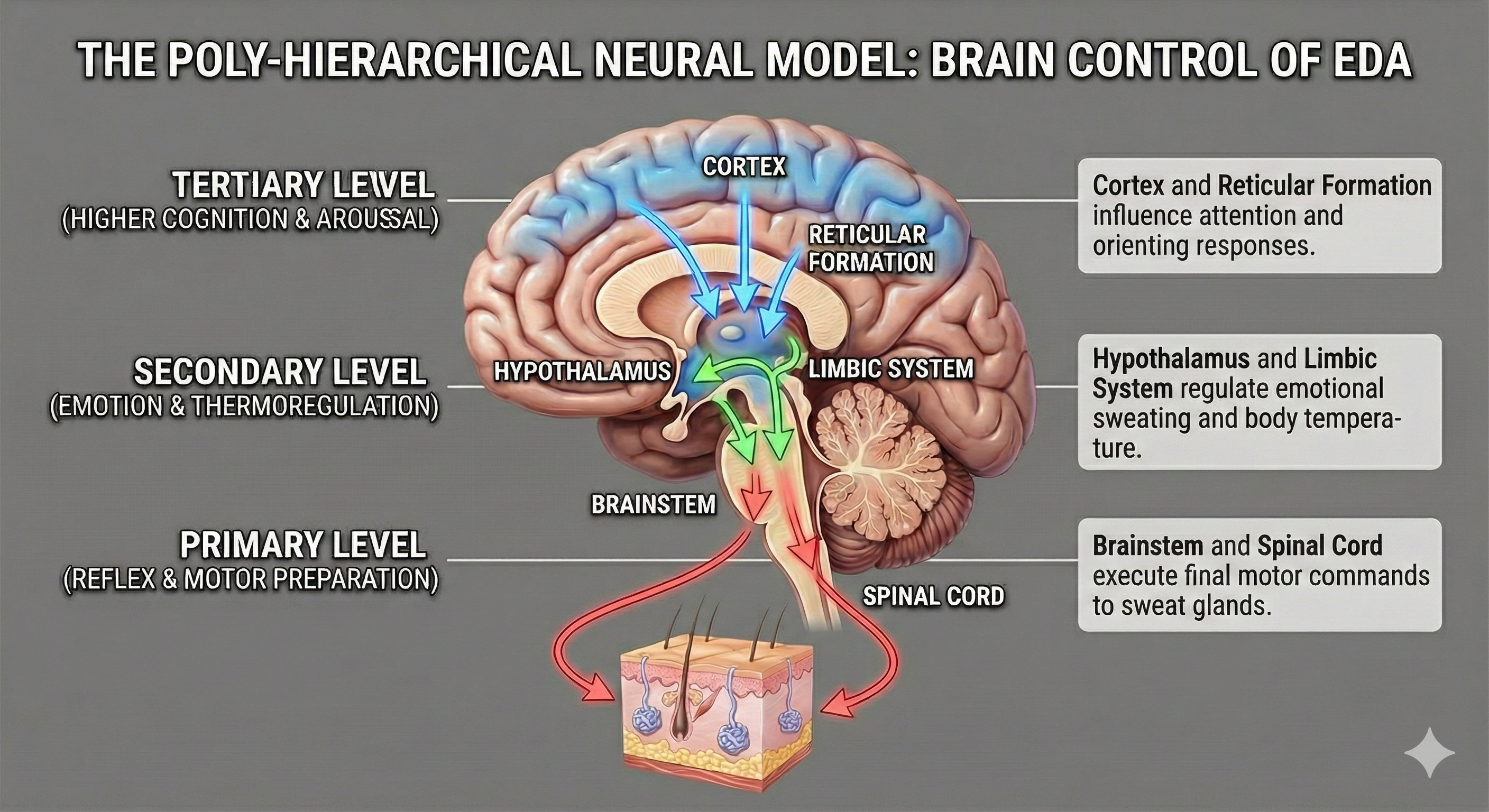
The Brain’s Command Center: A Three-Tiered Hierarchy
If the sweat glands are the hardware, the central nervous system provides the software. The poly-hierarchical neural model argues that EDA is not a single, monolithic "stress" signal but a composite of three distinct levels of brain activity. This hierarchy explains why we sweat for different reasons, such as heat, fear, or intense concentration.
The first and most primal level of control involves the hypothalamus and the amygdala. The hypothalamus acts as the body's thermostat, managing long-term, tonic shifts in skin moisture primarily for thermoregulation. However, because it is intimately connected to the amygdala—the brain’s fear center—this level is also responsible for the "emotional sweating" triggered by anxiety or threat detection (Amin & Faghih, 2022).
The second level involves the basal ganglia and the premotor cortex, areas of the brain dedicated to movement. Interestingly, EDA often spikes before a person physically reacts to a stimulus. The model suggests this is an evolutionary adaptation: the brain pre-moistens the palms to increase friction, improving our grip for "fight or flight" maneuvers.
The third and highest level of modulation comes from the reticular formation in the brainstem, which governs attention and arousal. When you encounter a novel stimulus—like a flashing light or a strange sound—the reticular formation generates a specific EDA spike known as the orienting response. This signal reflects the brain’s allocation of attention, effectively filtering what is important from background noise (Boucsein, 2012). By understanding these hierarchical levels, researchers can look at a squiggle on a graph and infer whether the subject is overheating, preparing to move, or simply paying attention.
This graphic illustrates the Poly-Hierarchical Neural Model, demonstrating how Electrodermal Activity (EDA) is regulated by a three-tiered brain system. It shows how higher cognitive and emotional centers (the Cortex, Reticular Formation, Hypothalamus, and Limbic System) influence the final motor commands executed by the Brainstem and Spinal Cord to activate sweat glands in the skin.
Eccrine Sweat Gland Function
Eccrine sweat glands have a secretory portion (coils) and a duct that opens on the skin surface. The sympathetic nervous system controls sweating through cholinergic fibers that release acetylcholine. Edelberg's poral valve model explains how the rapid opening and closing of sweat pores creates the characteristic shape of electrodermal responses.
Comprehension Questions: Sweat Gland Function and CNS Control
Why do sympathetic neurons release acetylcholine rather than norepinephrine at eccrine sweat glands?
How does Edelberg's Poral Valve model explain the rapid rise and fall of skin conductance during a response?
According to the Poly-Hierarchical Neural model, what three brain systems control electrodermal activity, and what function does each serve?
Factors That Influence EDA
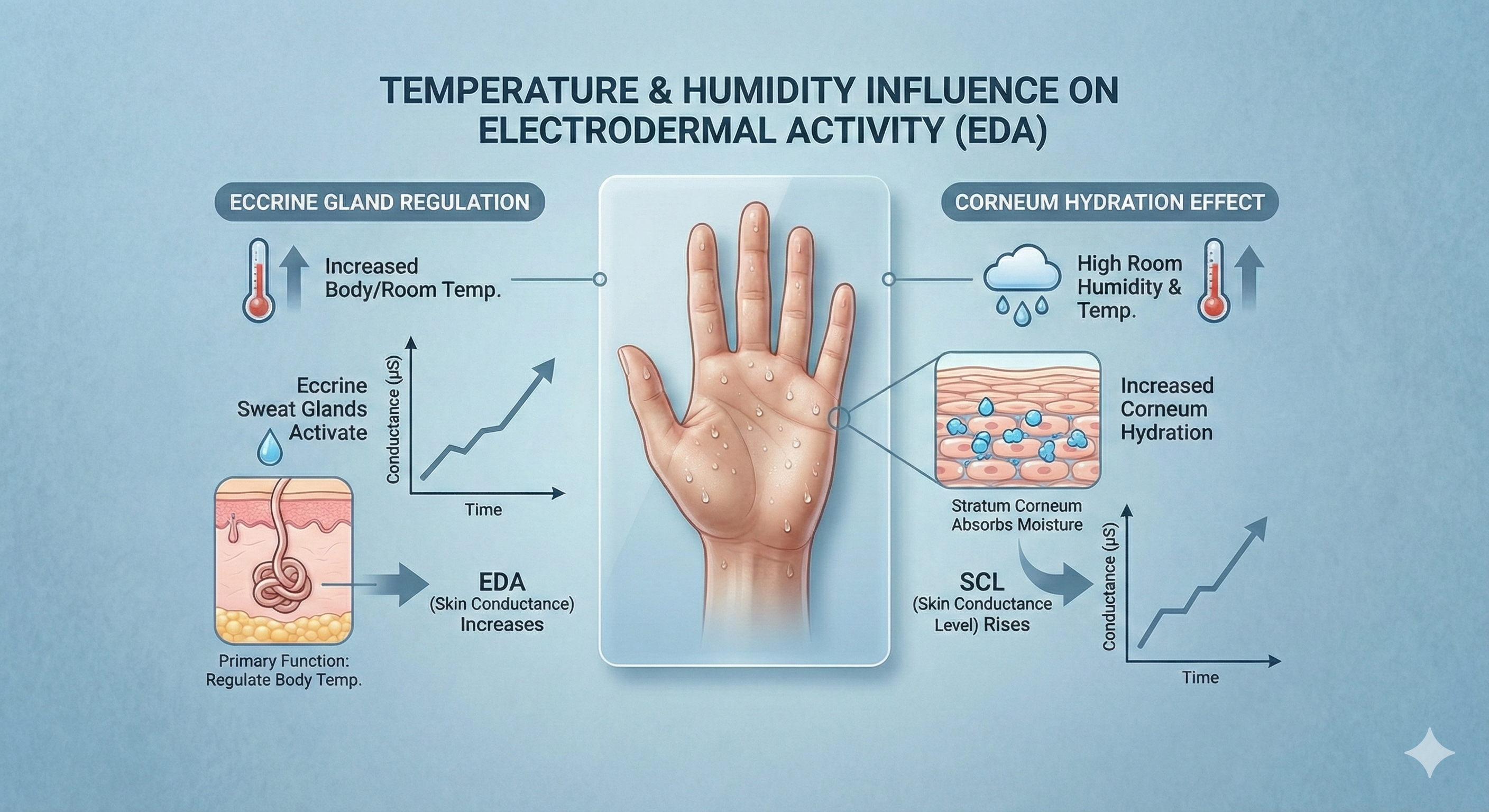
Temperature influences EDA since the primary function of eccrine sweat glands is to regulate body temperature. SCL rises with room humidity and temperature increases due to increased corneum hydration (Boucsein et al., 2012; Dawson et al., 2016).
The Environmental Influence on EDA: Visualizing how thermoregulation and corneal hydration drive changes in Skin Conductance Levels (SCL) in response to heat and humidity.
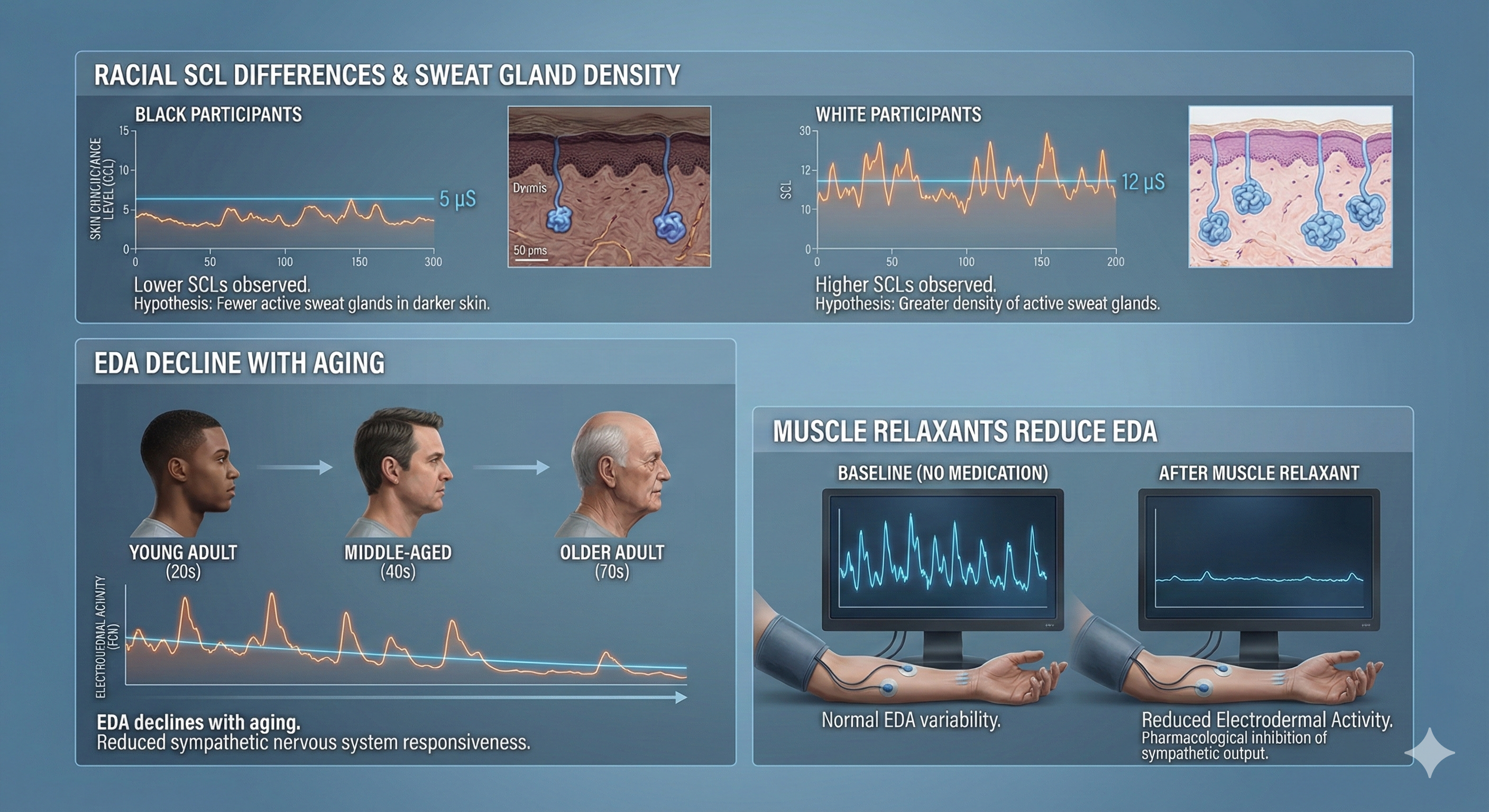
Gender differences in skin conductance level and reactivity may disappear when researchers control for a participant's most reactive hand (Roman et al., 1989). Boucsein (1992) reported lower SCLs for Black participants than their White counterparts. Racial SCL differences may be due to fewer sweat glands in dark skin. EDA declines with aging. Finally, muscle relaxants reduce electrodermal activity.
Factors Modulating Electrodermal Activity (EDA): An illustration showing the lower Skin Conductance Levels (SCL) observed in Black participants compared to White counterparts, the natural decline of EDA with aging, and the suppressive effects of muscle relaxants.
Controlling Environmental Variables
When you monitor electrodermal activity, remember that room temperature and humidity directly affect your readings. A client named Marcus comes in for his first biofeedback session on a hot summer day. His baseline SCL is significantly higher than expected. Before concluding that Marcus has elevated sympathetic arousal, consider whether the environmental conditions or his recent walk from the parking lot might explain the elevated readings. Always allow clients to acclimate to room temperature and document ambient conditions in your session notes.
Behavioral Correlates of EDA
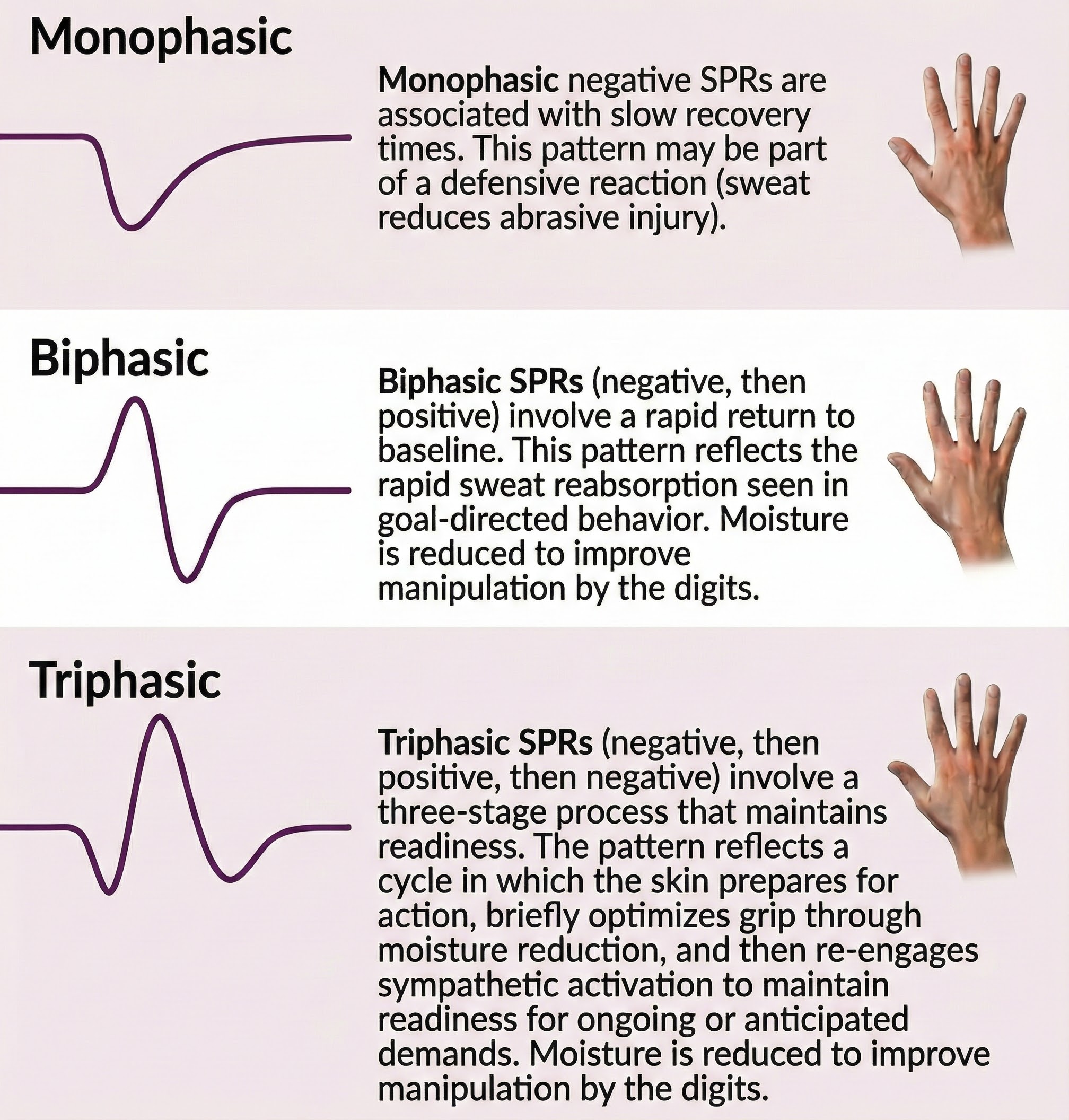
Skin potential response (SPR) can exhibit a monophasic negative or positive waveform, biphasic waveform (negative, then positive), or triphasic waveform (negative, positive, then negative). The classic pattern is biphasic. The illustration below was adapted from Stern, Ray, and Quigley (2001).
The functional significance and behavioral correlates of distinct Skin Potential Response (SPR) patterns.
Monophasic negative SPRs are associated with slow recovery times. This pattern may be part of a defensive reaction (sweat reduces abrasive injury).
Biphasic SPRs (negative, then positive) involve a rapid return to baseline. This pattern reflects the rapid sweat reabsorption seen in goal-directed behavior. Moisture is reduced to improve manipulation by the digits (Edelberg, 1970).
Triphasic SPRs (negative, then positive, then negative) involve an initial negative deflection, a return toward baseline, and then a secondary negative deflection. This pattern reflects a three-stage process: initial eccrine sweat gland activation during orienting or arousal, followed by sweat reabsorption characteristic of goal-directed behavior, and then renewed eccrine activation. The secondary negative wave suggests continued or renewed sympathetic arousal, possibly indicating sustained attention, anticipatory processing, or the preparation for subsequent action following an initial orienting response.
Does skin potential outperform skin conductance as a window into psychological states? The research says not really. Classic studies found that SP matched SC in sensitivity to stimulus intensity when looking at overall responses, though SP's multiple waveform components allowed researchers to detect finer gradations under certain scoring methods (Darrow, 1964; Burstein et al., 1965). Other work found substantial overlap between the two measures, with individual differences explaining much of the variation (Gaviria et al., 1969).
That said, SP does capture something different. Biophysical studies show that SP provides unique information about what is happening in the skin itself, including activity in the epidermis, the filling state of sweat ducts, and hydration levels, details that conductance alone cannot reveal (Edelberg, 1977; Tronstad et al., 2013; Pabst et al., 2017). When researchers combine SP with SC and susceptance measurements or use circuit-based models, they get a more complete picture of the skin's electrochemical state (Ehtiati et al., 2023).
So why does skin conductance dominate the field? Practicality. SC is easier to record, less finicky technically, and sensitive enough for most applications in emotion, arousal, and fear conditioning research. The extra information SP provides has not consistently translated into better prediction of psychological outcomes in large studies.
The bottom line: SP and related impedance measures add genuinely independent physiological dimensions beyond what conductance offers, particularly in waveform structure and timing. However, no study has yet shown that SP delivers a clear psychological advantage over SC across the board. The richest approach combines both measures rather than relying on either one alone.
The Old Question: Are You a "Steady" or "Reactive" Person?
Picture two friends watching a horror movie. One jumps at every scare, heart pounding, palms sweating. The other barely flinches. Are these two fundamentally different types of people, or is something more complex going on?
Scientists used to think they could sort people into categories: "labile" types who react strongly and change frequently, versus "stabile" types who stay calm and consistent. The research now shows this framing was too simple. Instead of being one type or the other, each of us carries both stable tendencies and changeable responses, and these operate somewhat independently.
What Sweaty Palms Taught Us
Early evidence came from measuring skin conductance, the subtle electrical changes that happen when your sweat glands activate slightly. Researchers found that people who showed lots of spontaneous skin conductance responses also tended to have higher baseline conductance and stronger orienting responses to new stimuli (Schell, Dawson, & Filion, 1988). This suggested a real dimension of physiological reactivity.
But here's the key insight: rather than discovering two distinct tribes of humans, researchers found a spectrum. Everyone has some baseline level of reactivity (a trait-like quality) plus moment-to-moment fluctuations (a state-like quality). Your sweaty palms during a job interview reflect both your general tendency toward physiological arousal and the specific stress of that situation.
Separating What Stays from What Shifts
Modern statistical methods called latent state-trait models let researchers mathematically separate these components (Steyer, Schmitt, & Eid, 1999; Steyer, Mayer, Geiser, & Cole, 2015). Think of it like this: if you measured someone's anxiety every day for a month, some portion of each score reflects their enduring anxious tendencies (the trait), while another portion reflects what happened that particular day (the state).
For major personality characteristics, the stable component typically accounts for most of the reliable variance, but the changeable component isn't trivial (Anusic & Schimmack, 2016; Bleidorn et al., 2022). People also become more consistent as they mature, with stability increasing through early adulthood before plateauing (Wagner, Lüdtke, & Robitzsch, 2019).
Why the Changeable Part Matters Most for Intervention
Here's where this gets practical. When researchers studied procrastination, they found something surprising: the fluctuating, state-like component of procrastination predicted students' study satisfaction better than the stable, trait-like component (Koppenborg, Ebert, & Klingsieck, 2024). The implication? Targeting the changeable part may yield bigger benefits than trying to overhaul someone's entire personality.
This principle applies broadly. Even vocational interests, which seem like fixed preferences, contain meaningful malleable variance (Roemer, Steinmayr, & Ziegler, 2023). And when researchers track people moment-to-moment, they find that individuals differ reliably not just in their average mood or behavior, but in how much they fluctuate (Jones, Brown, Serfass, & Sherman, 2017; Castro-Alvarez, Tendeiro, Meijer, & Bringmann, 2020).
What This Means for Biofeedback Practice
This trait-state framework has direct implications for biofeedback clinicians and clients.
Assessment becomes richer. When you conduct an initial psychophysiological stress profile, you're capturing a snapshot that reflects both trait-like baseline tendencies and state-like responses to that particular day and setting. A client who shows elevated muscle tension might have chronically high resting levels (a trait pattern), might be reacting to the novelty of the clinic (a state response), or both. Repeated assessments across sessions help you distinguish these components.
Training targets the state component. Biofeedback fundamentally works by teaching people to modify their state-level physiological responses. When a client learns to reduce skin conductance during a stressor, they're not erasing their trait-level reactivity; they're gaining voluntary control over state fluctuations. Over time, successful training may shift baseline levels too, but the immediate mechanism involves state regulation.
Individual differences in variability matter. Some clients show dramatic session-to-session swings in their physiological measures; others are remarkably consistent. Research suggests this variability itself is a meaningful individual difference (Castro-Alvarez et al., 2020). Highly variable clients may need different pacing, more frequent practice, or different feedback strategies than stable clients.
Expectations become realistic. Understanding the trait-state distinction helps set appropriate goals. A client with trait-level high sympathetic arousal won't become a naturally calm person, but they can learn to dampen their state-level spikes and recover more quickly. Framing success as "better regulation" rather than "personality transformation" aligns with what the science actually supports.
Transfer depends on state generalization. The ultimate goal of biofeedback is for skills to transfer outside the clinic. From a trait-state perspective, this means helping clients apply state-regulation strategies across diverse situations, gradually building a broader repertoire rather than changing a fixed trait.
The Bottom Line
You're not simply a "reactive" or "calm" person. You carry both stable tendencies and situation-specific responses, and biofeedback works primarily by giving you tools to regulate the changeable part. This framework explains why biofeedback can produce meaningful improvements without claiming to rewire someone's fundamental nature, and why consistent practice across varied conditions matters for lasting benefit.
Labiles show better control of overt emotional expression and are more responsive to environmental demands. Dean Drobot/Shutterstock.com.
These differences support the view that labiles better respond to changing environmental demands and allocate attentional resources to environmental events (Schell, Dawson, & Filion, 1988).
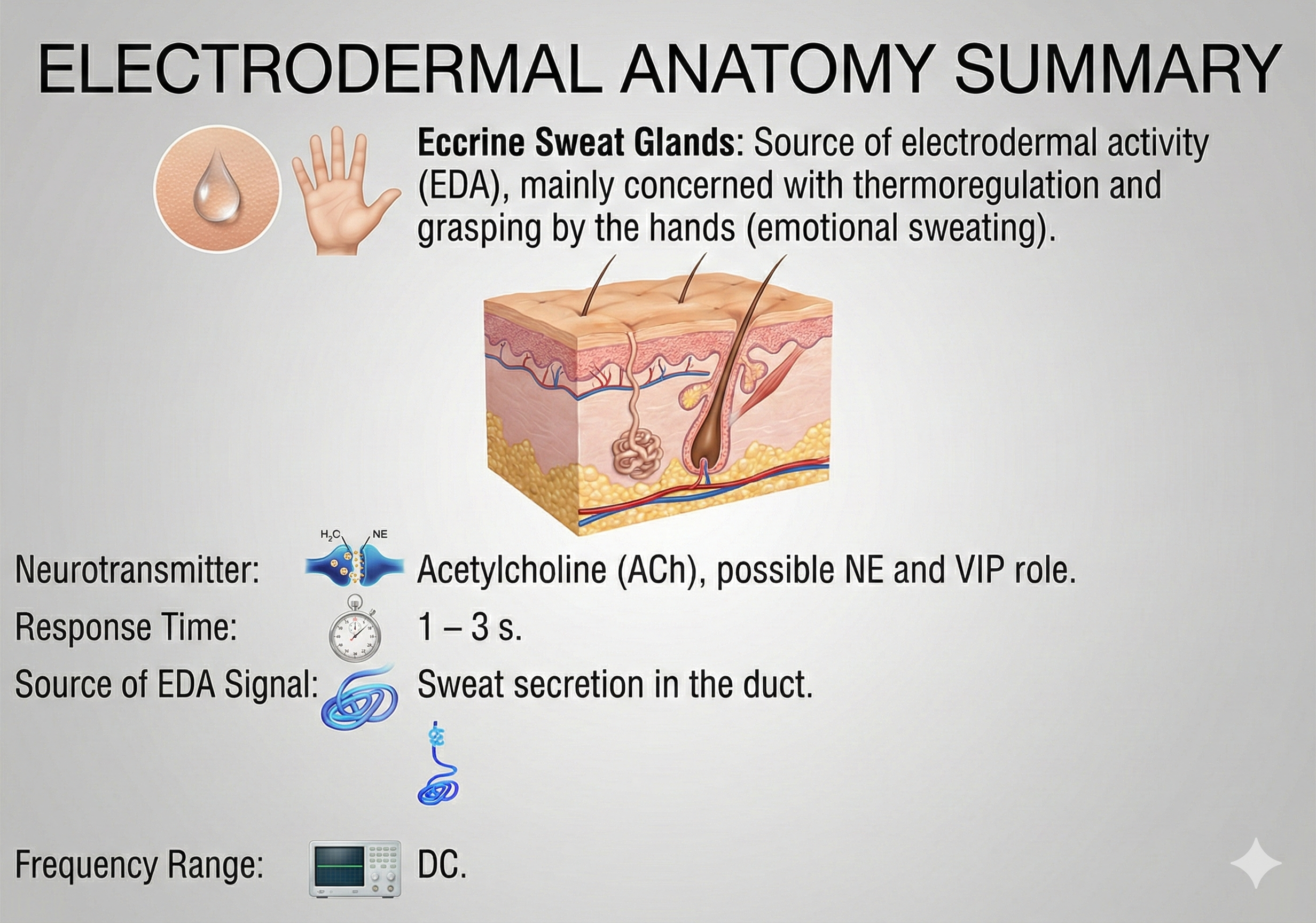
Summary of electrodermal anatomy concepts covered in this unit.
Behavioral Correlates and Individual Differences
SPR waveform patterns have behavioral significance: monophasic negative patterns suggest defensive reactions, while biphasic patterns indicate goal-directed behavior with rapid recovery. Individual differences in EDA define two types: labiles (higher SCL, larger SCR, faster responses, better vigilance) and stabiles (lower SCL, smaller SCR, slower responses). These differences reflect how individuals allocate attention and process information.
Comprehension Questions: Factors and Behavioral Correlates
How might environmental factors like room temperature affect your interpretation of a client's baseline SCL?
What does a biphasic SPR waveform suggest about the behavioral state of the person producing it?
How do labiles and stabiles differ in their electrodermal responses, and what might this mean for biofeedback training?
Why might skin potential provide different information than skin conductance?
A new client shows minimal SCRs during stress tasks. What individual difference factor might explain this pattern?
Cutting Edge Topics in Electrodermal Research
The following topics represent some of the most exciting research frontiers in electrodermal activity and biofeedback. These areas are still developing, with new studies appearing regularly, but they illustrate how our understanding of the electrodermal system continues to evolve.
Wearable EDA Monitoring: From the Lab to Real Life
Traditional electrodermal monitoring requires clients to sit still in a temperature-controlled room while sensors attached to their fingers detect changes in skin conductance. While this approach provides excellent signal quality, it tells us nothing about what happens when clients leave your office and face real-world stressors. The emergence of wearable EDA devices is changing this picture dramatically.
Modern wrist-worn devices can now continuously monitor electrodermal activity throughout the day, capturing responses to actual stressors rather than laboratory simulations. This ecological validity opens new possibilities for understanding how clients respond to their daily environments. Imagine tracking a client's EDA during a typical workday and identifying the specific situations that trigger their stress responses. This information could transform treatment planning by revealing patterns invisible during office-based assessment.
However, wearable EDA monitoring presents unique challenges. Movement artifacts, temperature fluctuations, and variable skin contact can all contaminate the signal. Researchers are developing sophisticated algorithms to separate genuine electrodermal responses from noise, but practitioners should interpret wearable data cautiously. The field is moving toward validation standards that will help clinicians know when wearable EDA data can be trusted for clinical decision-making.
Machine Learning and EDA Pattern Recognition
The human eye can recognize an obvious skin conductance response, but what about the subtle patterns embedded in continuous EDA recordings? Machine learning algorithms are increasingly capable of detecting meaningful patterns that escape visual inspection. These approaches can classify emotional states, predict anxiety episodes, and identify individual signatures in electrodermal responding.
One promising application involves using EDA patterns to predict the onset of anxiety or panic. If an algorithm could detect the early warning signs of an impending anxiety episode, it might trigger a just-in-time intervention, perhaps a prompt to practice breathing techniques delivered through a smartphone. This vision of responsive, automated biofeedback represents a significant departure from traditional session-based training.
Researchers are also exploring how machine learning can help identify the neural sources of electrodermal responses. By combining EDA recording with brain imaging and applying sophisticated analytical techniques, scientists hope to better understand how psychological states translate into peripheral physiological changes. This work may eventually help explain why some individuals show robust electrodermal responses while others remain relatively unreactive.
EDA and Interoceptive Awareness
Can you feel your skin conducting electricity? While the answer for most people is no, the relationship between electrodermal activity and body awareness is more complex than it might seem. Researchers are increasingly interested in how people perceive and interpret their own arousal states, and EDA provides a convenient window into this question.
Some studies suggest that individuals with anxiety disorders may have difficulty accurately perceiving their own physiological arousal. They might feel intensely anxious even when their skin conductance is relatively low, or fail to notice elevated arousal until it becomes overwhelming. This mismatch between physiological state and subjective experience may contribute to the maintenance of anxiety symptoms.
Biofeedback training may help by making normally invisible physiological signals visible and trackable. When clients can see their skin conductance on a screen, they gain an external reference point for calibrating their internal sense of arousal. Over time, this practice may improve interoceptive accuracy, helping clients develop a more accurate sense of their own physiological states without needing to look at a monitor. This perspective positions EDA biofeedback not just as a tool for reducing arousal, but as a method for improving body awareness.
Electrodermal Activity and Autonomic Flexibility
While heart rate variability has received enormous attention as an index of autonomic flexibility, electrodermal activity may tell a complementary story. Researchers are exploring whether patterns of EDA recovery, habituation, and response variability provide unique information about how flexibly the autonomic nervous system responds to changing demands.
The concept of electrodermal lability, discussed earlier in this unit, may be reconceptualized as an index of autonomic responsiveness. Rather than viewing labiles and stabiles as fixed personality types, current thinking emphasizes that electrodermal reactivity may shift based on context, health status, and training. This perspective suggests that biofeedback training might help stabiles develop greater electrodermal flexibility, potentially improving their ability to respond to environmental challenges.
Integration of EDA with other autonomic measures is an active research area. By simultaneously recording skin conductance, heart rate variability, and respiratory patterns, researchers hope to develop a more complete picture of autonomic regulation. This multimodal approach may reveal interactions between systems that are invisible when each measure is examined in isolation.
Glossary
acetylcholine: a neurotransmitter that nerve cells release to communicate with sweat glands and other target tissues; the primary chemical trigger for eccrine sweat production.
adrenergic receptors: cell surface proteins that respond to catecholamines such as adrenaline and noradrenaline.
apocrine sweat glands: glands that usually open into hair follicles and are mainly distributed in the armpits and genital region. They produce sweat odor, and distress can expel sweat from their tubules.
atropine: a drug that blocks muscarinic receptors and prevents acetylcholine from binding, effectively stopping sweat production.
biphasic SPR: a skin potential response (SPR) that starts with a negative phase and then ends with a positive phase.
brainstem reticular formation: the lowest level of central EDA control that initiates electrodermal activity when we encounter novel or important stimuli.
contralateral system: in Boucsein's model of EDA control, a system that controls the hand's sweat gland activity on the opposite side of the brain. This system expresses activity in the lateral prefrontal cortex and the basal ganglia (i.e., caudate nucleus and putamen), and mediates EDA during cognition, orienting, and locomotion.
catecholamines: stress hormones including adrenaline and noradrenaline that can influence sweating intensity, particularly during intense emotional experiences.
co-transmitters: signaling molecules released alongside the primary neurotransmitter to modify or enhance its effects.
dermis: the inner skin layer that contains blood and lymph vessels, smooth muscle, and sebaceous and sweat glands.
eccrine sweat glands: glands that respond to cognitive activity, emotion, and temperature, achieve thermoregulation (temperature control) through evaporative cooling and are responsible for EDA.
electrodermal activity (EDA): changes in the electrical properties of the skin resulting from sweat gland activity controlled by the sympathetic nervous system.
emotional sweating: sweating that begins at the palms, soles, and armpits in response to fear or embarrassment.
endosomatic: from within the body. Skin potential is measured by detecting voltage differences between two electrodes on the skin surface.
epidermis: outer skin layer that consists of five layers (stratum corneum, stratum lucidum, stratum granulosum, stratum spinosum, and stratum germinativum).
exocrine glands: glands that secrete into ducts.
exosomatic: from outside the body. Skin conductance and skin resistance are measured by passing an electric current through the skin.
galvanic skin response (GSR): an older term for skin resistance, reflecting opposition to external current movement.
hypodermis: the region below the dermis that consists of connective tissue and contains the secretory portion of sweat glands and blood and lymph vessels.
ipsilateral system: in Boucsein's model of EDA control, a system that controls the sweat gland activity of the hand on the same side of the brain.
ion channels: protein tunnels embedded in cell membranes that allow charged particles to move across the membrane.
labiles: individuals who show higher resting SCLs and larger SCRs, and more rapid responses to stimuli and return to resting levels, less overt emotional expression, and more agreeableness than stabiles. Labiles may better respond to changing environmental demands and allocate attentional resources to environmental events.
level: a tonic measure of EDA that quantifies the average amplitude over a specified period.
locomotor system: in Hugdahl's model of EDA control, this system consists of the premotor cortex, pyramidal tract, and brainstem. The locomotor system hydrates the soles of the feet and palms of the hands to increase running speed and hand dexterity.
lumen: the hollow center of a gland or tubular structure where secretions accumulate before being released.
Malpighian layer: the deepest layer of the epidermis. Sweat glands are typically filled to this layer.
monophasic negative SPRs: skin potential response (SPR) with one negative limb.
myoepithelial cells: the cells that line the coils of the secretory portion of a sweat gland. Myoepithelial cells produce sweat and may contribute to spontaneous EDA.
muscarinic M3 receptors: specialized proteins embedded in cell membranes that recognize and respond specifically to acetylcholine, triggering sweat production when activated.
neuropeptides: small protein-like molecules used by neurons to communicate, contributing to intrinsic regulation of gland activity.
nitric oxide: a small gaseous signaling molecule produced within tissues that contributes to local regulation of physiological processes.
norepinephrine: a neurotransmitter typically associated with arousal and alertness, commonly released by sympathetic nerve endings throughout most of the body but notably absent at eccrine sweat glands.
orienting-arousal system: in Hugdahl's model of EDA control, this system consists of the lateral frontal cortex, amygdala and hippocampus, and reticular formation. The orienting-arousal system produces sweating to protect the skin from injury in situations demanding focused attention (e.g., novel stimuli) or vigilance for threats.
phasic: a brief change in physiological activity in response to a discrete stimulus. For example, a single skin potential response in reaction to a sudden tone.
peptide histidine isoleucine: a co-transmitter peptide found in nerve fibers surrounding eccrine glands that helps regulate gland activity and local blood flow.
poly-hierarchical neural model: the integrated model that proposes that electrodermal activity is regulated by multiple parallel control systems operating at different levels of the nervous system, including spinal, brainstem, limbic, and cortical structures, each capable of independently modulating sweat gland activity while also interacting with one another to produce integrated autonomic responses.
poral valve model: Edelberg's revised model of a skin conductance response (SCR) in which rising levels of sweat in the duct open a poral valve, depositing sweat on the skin surface.
preoptic area of the hypothalamus: forebrain structure that receives reports of body temperature and activates sympathetic nerves that innervate eccrine sweat glands to initiate thermoregulation through evaporative cooling.
resistance: the reciprocal of conductance, also called galvanic skin response (GSR), that reflects opposition to external current movement and is measured as skin resistance level (SRL) and skin resistance response (SRR) in Kohms (thousands of ohms).
resistor: an electrical or electronic component that opposes electrical current flow. Edelberg described the long, tubular sweat ducts as resistors wired in parallel.
response: a phasic measure of EDA representing a spontaneous or stimulus-elicited change in sweat gland activity.
secretory portion: the coils of an eccrine sweat gland that produce sweat.
skin conductance: an exosomatic measure of EDA that indexes how easily an external current passes through the skin that is measured as skin conductance level (SCL) and skin conductance response (SCR) in microsiemens (formerly micromhos).
skin conductance level (SCL): a tonic measure of skin conductance measured in microsiemens.
skin conductance response (SCR): a phasic measure of skin conductance measured in microsiemens.
skin potential: an endosomatic measure of EDA that detects voltage differences between two electrodes on the skin surface. Skin potential is measured as skin potential level (SPL) and skin potential response (SPR) in millivolts (thousandths of a volt).
skin potential level (SPL): a tonic measure of skin potential measured in millivolts.
skin potential response (SPR): a phasic measure of skin potential measured in millivolts.
skin resistance: an exosomatic measure of EDA that reflects opposition to current movement and is measured as skin resistance level (SRL) and skin resistance response (SRR), also called galvanic skin response (GSR), in Kohms (thousands of ohms).
skin resistance level (SRL): a tonic measure of skin resistance measured in Kohms.
skin resistance response (SRR): a phasic measure of skin resistance measured in Kohms.
stabiles: individuals who show lower resting SCLs and smaller SCRs, slower responses to stimuli and return to resting levels, more overtly emotionally expressive, and more antagonistic than labiles. They seem less effective than labiles in responding to changing environmental demands and allocating attentional resources to environmental events.
sudoriferous glands: apocrine and eccrine sweat glands.
sweat circuit model: Edelberg's hypothesis that EDA is a function of sweat duct filling and action by a selective membrane in the epidermis. He proposed that duct-filling produces SCRs, while both duct-filling and the selective membrane control response recovery.
sympathetic cholinergic fibers: unmyelinated sympathetic motoneurons that mainly innervate eccrine sweat glands. These motoneurons release ACh at the secretory portion. A few may target the sweat duct.
sympathetic nervous system: autonomic nervous system branch that regulates activities that expend stored energy and primarily controls EDA.
thermoregulation: temperature control through evaporative cooling.
thermoregulatory system: in Hugdahl's model of EDA control, this system lies in the anterior hypothalamus and produces cold sweating during trauma. Increased sweating is accompanied by constriction of peripheral blood vessels and increased EDA in the hands and digits.
tonic: the background level of physiological activity. For example, a 5-minute average of skin conductance activity.
transporters: proteins that actively shuttle substances from one side of a cell membrane to the other.
vasoactive intestinal peptide (VIP): a co-transmitter peptide found in nerve fibers around eccrine glands that binds to secretory cells and helps regulate gland activity and local blood flow.
Test Yourself
Click the button below to take 10-question tests over this unit without an exam password.
BioSource Software offers Human Physiology, which satisfies BCIA's Human Anatomy and Physiology requirement, and Biofeedback100, which provides extensive multiple-choice testing over BCIA's Biofeedback Blueprint.
Assignment
Now that you have completed this module, describe the difference between level and response, exosomatic and endosomatic measurements, and labiles and stabiles. Summarize the main reasons for palmar sweating.
References
Andreassi, J. L. (2007). Psychophysiology: Human behavior and physiological response (5th ed.). Lawrence Erlbaum and Associates, Inc.
Bach, D. R. (2014). Sympathetic nerve activity can be estimated from skin conductance responses: A comment on Henderson et al. (2012). NeuroImage, 84, 122-123. https://doi.org/10.1016/j.neuroimage.2013.08.030
Banks, S. J., Bellerose, J., Douglas, D., & Jones-Gotman, M. (2012). Bilateral skin conductance responses to emotional faces. Applied Psychophysiology and Biofeedback, 37(3), 145-152. https://doi.org/10.1007/s10484-011-9177-7
Boucsein, W. (1992). Electrodermal activity. Plenum Press.
Boucsein, W., & Hoffmann, G. (1979). A direct comparison of the skin conductance and skin resistance methods. Psychophysiology, 16(4), 316–322. https://doi.org/10.1111/j.1469-8986.1979.tb01488.x
Boucsein, W., Fowles, D. C., Grimnes, S., Ben-Shakhar, G., Roth, W. T., Dawson, M. E., & Filion, D. L. (2012). Publication recommendations for electrodermal measurements. Psychophysiology, 49(8), 1017-1034. https://doi.org/10.1111/j.1469-8986.2012.01384.x
Breedlove, S. M., & Watson, N. V. (2020). Behavioral neuroscience (9th ed.). Sinauer Associates.
Burstein, K. R., Fenz, W., Bergeron, J., & Epstein, S. (1965). A comparison of skin potential and skin resistance responses as measures of emotional responsivity. Psychophysiology, 1(1), 48–62. https://doi.org/10.1111/j.1469-8986.1964.tb02607.x
Cui, C., & Schlessinger, D. (2015). Eccrine sweat gland development and sweat secretion. Experimental Dermatology, 24(9), 644–650. https://doi.org/10.1111/exd.12757
Darrow, C. W. (1964). The rationale for treating the change in galvanic skin response as a change in conductance. Psychophysiology, 1(1), 31–38. https://doi.org/10.1111/j.1469-8986.1964.tb02606.x
Davidson, R. J., Ekman, P., Saron, C. D., Senulis, J. A., & Friesen, W. V. (1992). Approach-withdrawal and cerebral asymmetry: Emotional expression and brain physiology I. Journal of Personality and Social Psychology, 58(2), 330-341.
Dawson, M. E., Schell, A. M., & Filion, D. L. (2016). The electrodermal system. In J. T. Cacioppo, L. G. Tassinary, & G. G. Berntson (Eds.), Handbook of psychophysiology (4th ed.). Cambridge University Press.
Edelberg, R. (1970). The information content of the recovery limb of the electrodermal response. Psychophysiology, 6(5), 527-539.
Edelberg, R. (1972). Electrical activity of the skin: Its measurement and uses in psychophysiology. In N. S. Greenfield & R. A. Sternbach (Eds.), Handbook of psychophysiology (pp. 367-418). Holt, Rinehart & Winston.
Edelberg, R. (1973). Mechanisms of electrodermal adaptations for locomotion, manipulation, or defense. Progress in Physiological Psychology, 5, 155-209.
Edelberg, R. (1977). Relation of electrical properties of skin to structure and physiologic state. Journal of Investigative Dermatology, 69(3), 324–327. https://doi.org/10.1111/1523-1747.ep12498361
Edelberg, R. (1993). Electrodermal mechanism: A critique of the two-effector hypothesis and a proposed replacement. In J. C. Roy, W. Boucsein, D. C. Fowles, & J. H. Gruzelier (Eds.), Progress in electrodermal research (pp. 7-30). Plenum Press.
Eedy, D., Johnston, C., Shaw, C., & Buchanan, K. (1990). Peptidergic neurotransmitter innervation of the eccrine sweat gland and their presence in sweat secretions. British Journal of Dermatology, 123(1), 103–109. https://doi.org/10.1111/j.1365-2133.1990.tb06042.x
Ehtiati, K., Eiler, J., Bochyńska, A., Nissen, L. L., Strøbech, E., Nielsen, L. F., & Thormann, E. (2023). Skin and artificial skin models in electrical sensing applications. ACS Applied Bio Materials, 6(8), 3224–3248. https://doi.org/10.1021/acsabm.3c00298
Fowles, D. C. (1986). The eccrine system and electrodermal activity. In M. G. H. Coles, E. Donchin, & S. W. Porges (Eds.), Psychophysiology, systems, processes & applications (pp. 51-96). Guilford.
Freeman, R., Waldorf, H. A., & Dover, J. S. (1992). Autonomic neurodermatology (Part II): Disorders of sweating and flushing. Seminars in Neurology, 12(4), 395–405. https://doi.org/10.1055/s-2008-1041212
Gaviria, B., Coyne, L., & Thetford, P. E. (1969). Correlation of skin potential and skin resistance measures. Psychophysiology, 5(2), 153–167. https://doi.org/10.1111/j.1469-8986.1968.tb02838.x
Hugdahl, K. (1995). Psychophysiology: The mind-body perspective. Harvard University Press.
Harker, M. (2013). Psychological sweating: A systematic review focused on aetiology and cutaneous response. Skin Pharmacology and Physiology, 26(2), 92–100. https://doi.org/10.1159/000346930
Hu, Y., Converse, C., Lyons, M. C., & Hsu, W. (2018). Neural control of sweat secretion: A review. British Journal of Dermatology, 178(6), 1246–1256. https://doi.org/10.1111/bjd.16156
Jacob, S. W., & Francone, C. A. (1970). Structure and function in man. Saunders.
Katkin, E. S. (1975). Electrodermal lability: A psychophysiological analysis of individual differences in response to stress. In I. G. Sarason & C. D. Spielberger (Eds.), Stress and anxiety (Vol. 2, pp. 141-176). Wiley.
Klaka, P., Grüdl, S., Banowski, B., Giesen, M., Sättler, A., Proksch, P., Welss, T., & Förster, T. (2017). A novel organotypic 3D sweat gland model with physiological functionality. PLOS ONE, 12(8), e0182752. https://doi.org/10.1371/journal.pone.0182752
LaBar, K. S., Gatenby, J. C., Gore, J. C., LeDoux, J. E., & Phelps, E. A. (1998). Human amygdala activation during conditioned fear acquisition and extinction: A mixed-trial fMRI study. Neuron, 20, 937-945. https://doi.org/10.1016/s0896-6273(00)80475-4
Lacey, J. I., & Lacey, B. C. (1958). The relationship of resting autonomic activity to motor impulsivity. In J. I. Lacey & B. C. Lacey (Eds.), The brain and human behavior (pp. 144-209). Williams & Wilkins.
Machado-Moreira, C. A., McLennan, P. L., Lillioja, S., van Dijk, W., Caldwell, J. N., & Taylor, N. A. (2012). The cholinergic blockade of both thermally and non-thermally induced human eccrine sweating. Experimental Physiology, 97(5), 544–555. https://doi.org/10.1113/expphysiol.2011.062984
Pabst, O., Tronstad, C., Grimnes, S., Fowles, D. C., & Martinsen, Ø. G. (2017). Comparison between the AC and DC measurement of electrodermal activity. Psychophysiology, 54(3), 374–385. https://doi.org/10.1111/psyp.12788
Roman, R. J., Sevastos, P. P., & Thomas, S. A. (1989). Sex differences in electrodermal lability. Psychophysiology, 26(S4A), S54.
Schell, A. M., Dawson, M. E., & Filion, D. L. (1988). Psychophysiological correlates of electrodermal lability. Psychophysiology, 25(6), 619-632.
Schwartz, M. S., & Andrasik, F. (Eds.). (2003). Biofeedback: A practitioner's guide (3rd ed.). The Guilford Press.
Shaffer, F., Combatalade, D., Peper, E., & Meehan, Z. (2016). A guide to cleaner electrodermal activity measurements. Biofeedback, 44(2), 90-100. https://doi.org/10.5298/1081-5937-44.2.01
Shields, S. A., MacDowell, K. A., Fairchild, S. B., & Campbell, M. L. (1987). Is mediation of sweating cholinergic, adrenergic, or both? A comment on the literature. Psychophysiology, 24(3), 312-319. https://doi.org/10.1111/j.1469-8986.1987.tb00301.x
Stern, R. M., Ray, W. J., & Quigley, K. S. (2001). Psychophysiological recording (2nd ed.). Oxford University Press.
Tortora, G. J., & Derrickson, B. H. (2021). Principles of anatomy and physiology (16th ed.). John Wiley & Sons, Inc.
Tronstad, C., Kalvøy, H., Grimnes, S., & Martinsen, Ø. G. (2013). Waveform difference between skin conductance and skin potential responses in relation to electrical and evaporative properties of skin. Psychophysiology, 50(10), 920–927. https://doi.org/10.1111/psyp.12073
Venables, P. H., & Martin, I. (1967). The relation of palmar sweat gland activity to level of skin potential and conductance. Psychophysiology, 3(3), 302-311.
Wallin, B. G. (1981). Sympathetic nerve activity underlying electrodermal and cardiovascular reactions in man. Psychophysiology, 18(4), 470-476.
Wilson, J. (2003). Biological foundations of human behavior. Wadsworth/Thompson Learning.
Zancanaro, C., Merigo, F., Crescimanno, C., Orlandini, S., & Osculati, A. (1999). Immunohistochemical evidence suggests intrinsic regulatory activity of human eccrine sweat glands. Journal of Anatomy, 194(3), 401–408. https://doi.org/10.1046/j.1469-7580.1999.19430401.x